2017.01.21 11:03:42
Таблица митохондриального композиционного генетического кода
http://www.uniprot.org/uniprot/P68530
http://www.ebi.ac.uk/ena/data/view/CAA24000
Вторичная структура (Пикотех-2017): https://img-fotki.yandex.ru/get/104083/ … 0_orig.png
{kind=link}

Подобная вторичная структура является косвенным подтверждением митохондриального кода триптофана "3"

А таблица митохондриального композиционного генетического кода будет выглядеть так:


Наличие типовых структем, например, программных спиралей, является дополнительным косвенным подтверждением существования митохондриального композиционного генетического кода.
http://www.uniprot.org/uniprot/P00415
http://www.ebi.ac.uk/ena/data/view/AAQ06599
Вторичная структура (Пикотех-2017): https://img-fotki.yandex.ru/get/104083/ … c_orig.png
{kind=link}

Структемы-перевертыши убеждают в том, что митохондриальный композиционный генетический код возник на ранних этапах эволюции генетического аппарата, т.е. "на заре белкового мира".
http://www.uniprot.org/uniprot/P00423
http://www.ebi.ac.uk/ena/data/view/AAA30461
Вторичная структура (Пикотех-2017): https://img-fotki.yandex.ru/get/104083/ … 7_orig.png
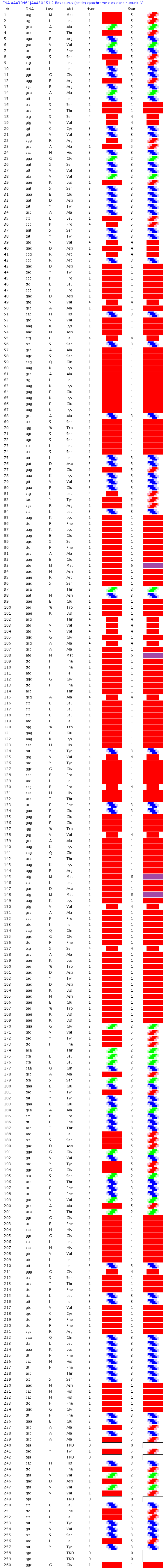{kind=link}

Коды "4" скручивают альфа-спираль, а коды "6" наклоняют. Таким образом композиционный генетический код управляет ростом альфа-310-Met-спирали по двум степеням свободы.
http://www.uniprot.org/uniprot/P00426
http://www.ebi.ac.uk/ena/data/view/AAS20602
>ENA|AAS20602|AAS20602.1 Bos taurus (cattle) mitochondrial cytochrome c oxidase subunit 5a
ATGCTAGGCGCCGCTGTCCGCCGCTGCTCTGTAGCTGCAGCCGCAGTCGCCCGGGCCAGC
CCTCGAGGCCTCCTGCACCCCACTCCGGCCCCCGGCCAAGCCGCCGCTGTCCAGTCACTT
CGCTGCTACTCCCATGGGTCACATGAGACAGACGAGGAGTTTGATGCTCGCTGGGTGACA
TACTTCAATAAGCCAGATATTGATGCTTGGGAGTTGCGTAAAGGGATGAACACACTTGTT
GGCTATGACCTGGTTCCAGAGCCCAAAATCATTGATGCTGCTTTGCGGGCATGCAGACGT
TTAAATGATTTTGCTAGTGCAGTTCGCATTCTAGAAGTTGTTAAGGACAAAGCAGGACCT
CATAAGGAAATCTACCCCTATGTCATCCAGGAACTTAGACCAACTTTGAATGAACTGGGA
ATCTCCACTCCAGAGGAACTGGGCCTTGACAAAGTGTAA
Вторичная структура (Пикотех-2017): https://img-fotki.yandex.ru/get/194425/ … c_orig.png
{kind=link}
2017.01.22 16:18:50
Пикотехнология белков, ДНК, РНК
Музыкальные конвейеры белкового мира
Каждый активный центр белка может представлять молекулярного "оператора", который дистанционно или в непосредственном контакте работает с группами атомов или отдельными ионами / электронами. Последовательность активных центров напоминает сборочный конвейер, где из деталей (элементарных частиц) и блоков разного уровня (атомов, молекул...) собираются рабочие пикомашины. Некоторые белковые молекулы или их группы образуют не один конвейер, а разветвлённую конвейерную сеть, где есть много входных и выходных портов. Каждая такая сеть может быть собрана за доли секунды и "жить" десятки минут. Новая версия программы Пикотехнология поможет увидеть структуры и понять нюансы работы этих конвейерных сетей белкового мира.



Типовой блок (аминокислота) для сборки пикомашины, которая может оказаться очередной конвейерной сетью...
Как работает "оператор"(активный центр белка) я уже писал в рассылке. Гармоничные сочетания гиперзвуков могут дистанционно ориентировать и двигать микрообъекты по любому направлению. Таким образом осуществляется правильная стыковка исходных объектов и передача от "оператора" к "оператору" вдоль конвейера или по конвейерной сети.

Примерно так же работает сравнительно простой белковый конвейер-молекула. Только на несколько порядков быстрее и надёжнее ![]()
[11:48:24] Кушелев Александр Юрьевич: Сейчас создаётся новая 3D версия, в которой будут адекватно учтены углы скручивания и изгиба спиралей. А 2D-версия уже готова:

[11:49:33] Andrei: а док-во вы отказываетесь искать
[11:49:36] Кушелев Александр Юрьевич: Заказчик / заказчица может убедиться в полезности 2D-версии программы Пикотех. Даже 2D-версия может дать больше полезной информации, чем 3D-модели из PDB
[11:49:54] Кушелев Александр Юрьевич: Заказчики сами разберутся, нужна им информация или нет.
[11:50:05] Кушелев Александр Юрьевич: А Вы им её не даёте, поэтому у них и интерес пропал.
[11:50:52] Кушелев Александр Юрьевич: Объясните, что мы работаем на уровне пикометров:

[11:52:17] Кушелев Александр Юрьевич: И та информация, которую даёт даже 2D-Пикотех, может оказаться гораздо полезнее информации из PDB. Тем более, что "лишней информации не бывает". Мы же даём дополнительную информацию
[11:53:01] Andrei: вы опять за свои "сказки" принялись бездоказательные
[11:53:27] Andrei: это называется синдром изобретателя
[11:53:40] Andrei: им мерещаться гениальные открытия
[11:53:55] Кушелев Александр Юрьевич: Чтобы понять научную ценность, нужно владеть материалом. Вы кто по образованию?
[11:54:38] Andrei: а хоть один с образованием кого вы встречали ваши сказания может подтвердить?
[11:54:55] Кушелев Александр Юрьевич: Вы с какой Луны упали?
[11:55:06] Кушелев Александр Юрьевич: Виктория Соколик по нашим материалам книгу написала
[11:55:56] Andrei: и что это доказывает?
[11:55:58] Кушелев Александр Юрьевич: Одна лишь таблица композиционного генетического кода может дать больше информации, чем вся база PDB
[11:56:10] Andrei: опять 25
[11:56:12] Кушелев Александр Юрьевич: 64 цифры
[11:56:29] Andrei: все это пока гипотеза бездоказательная
[11:57:09] Andrei: вы даже не осилили статистический анализ сделать по научному между вашей программой и Базой
[11:57:39] Кушелев Александр Юрьевич: Каждый потенциальный заказчик сам может решить, что доказано, что не доказано. Вы-то кто по образованию? Кристалл от молекулы сможете отличить?
[11:58:11] Andrei: речь о том можете ли вы решить сами или нет
[11:58:21] Кушелев Александр Юрьевич: Вы за форумом следите?
[11:58:32] Andrei: ибо доказательств у вас ноль
[11:59:12] Andrei: и полосочки цветные не сложно сгенирировать на компе
[11:59:20] Кушелев Александр Юрьевич: aest просил 6 фрагментов, замкнутых через дисульфидные мостики. Вот они
[11:59:52] Кушелев Александр Юрьевич: А Вы про полосочки... Вы же молекулу от кристалла не отличите
[12:00:24] Andrei: все бы хорошо, но доказаетельств что ваши полосочки правильны на самом деле у вас нет
[12:00:37] Кушелев Александр Юрьевич: Про отличие молекулы от атома я Вас уже не спрашиваю. Это всё равно, что от козла молока ждать
[12:00:59] Andrei: так это я от вас жду молока но безрезультатно
[12:01:01] Кушелев Александр Юрьевич: Вы взялись за работу менеджера, а сами лезете в науку.
[12:01:21] Кушелев Александр Юрьевич: Зачем мне такой "менеджер"?
[12:01:45] Andrei: приходится лезть когда изобретатель безграмотный в методах научных доказательств
[12:01:45] Кушелев Александр Юрьевич: Если Вы сами в науке разбираетесь, то я Вам не нужен
[12:02:06] Кушелев Александр Юрьевич: Прощайте. С Вами только время терять
[12:02:48] Andrei: есть сейчас один эксперт можно попробовать ее спросить
[12:03:22] Andrei: но если вы будете вести себя неадекватно то смысла нет этим заниматься вообще
[12:24:19] Andrei:
Кушелев: заказчик / заказчица может убедиться в полезности 2D-версии программы Пикотех. Даже 2D-версия может дать больше полезной информации, чем 3D-модели из PDB
Андрей: там и так была 2D модель
[12:24:40] Andrei: смысла заново слать нет
[12:31:14] Кушелев Александр Юрьевич: Вы мне писали, что послали только 3D-модели
[12:31:49] Andrei: все что было то и послал
[12:32:03] Andrei: там и 2Д тоже была
[12:32:44] Кушелев Александр Юрьевич: Это меняет дело
[12:32:45] Andrei: но в любом случае это не заинтерсовало из-за ошибки в 6 раз от РСА
[12:33:18] Кушелев Александр Юрьевич: Надо было объяснить, что 3D версия ещё не протестирована. Обнаружились ошибки. Зато 2D-версия работает безошибочно
[12:33:36] Andrei: так еще хуже будет
[12:33:44] Кушелев Александр Юрьевич: Ну и заодно объяснить, что точность моделей в PDB ещё хуже
[12:33:55] Andrei: надо молчать пока никто не понимает как оно на самом деле
[12:34:18] Andrei: там хоть фигу рисуй никто не сможет ни доказать ни опровергнуть
[12:34:22] Кушелев Александр Юрьевич: Лучше открыть глаза заказчикам на реальную погрешность моделей из PDB
[12:34:34] Кушелев Александр Юрьевич: Иначе они на PDB молятся, а там пшик...
[12:35:10] Andrei: вот как докинг сделаете автоматический то тогда другое дело
[12:35:30] Andrei: а пока там можно фиги рисовать и никто не удивится
[12:35:41] Кушелев Александр Юрьевич: Тогда ждите. А другие менеджеры будут работать
[12:35:53] Andrei: я только за
[12:37:03] Andrei: когда хоть одному удасться что-то продать то пишите
[12:37:13] Andrei: это может изменить положение дел
[12:38:26] Кушелев Александр Юрьевич: Конечно изменит! И 80% тоже изменятся
[12:39:09] Andrei: лучше 1% от милиона чем 80% от нуля
[12:40:09] Кушелев Александр Юрьевич: Поэтому Вы до сих пор и на нуле
[12:40:40] Andrei: ноль это то что имеем от изобретателя на данный момент
[12:41:21] Andrei: тут хоть 800% пообещайте и будет то же самое
[12:41:35] Кушелев Александр Юрьевич: Мои модели уже школьники с 2004 года изучают, а Вы в 2017-ом году ещё тормозите...
[12:42:13] Andrei: Ну ок... ждем результатов от новых менеджеров нетормозных
[12:43:20] Кушелев Александр Юрьевич: Благодарю за то, что нашли время для нашей научной дискуссии
[12:44:31] Andrei: но если и новые менеджеры не осилят ничего то нужно будет серьезно научно задуматься "а есть ли ребенок"
[13:17:02] Andrei: https://www.mathworks.com/help/bioinfo/ … hworks.com
[13:18:57] Andrei: https://www.mathworks.com/help/bioinfo/ … cules.html
[13:21:29] Кушелев Александр Юрьевич: Когда напишите, чем отличается кристалл от молекулы, тогда продолжим научную дискуссию ![]()
[13:36:01] Andrei: вы подозрительно невнимательны, там речь идет об атомах имеющих кольцегранную структуру
[13:36:37] Andrei: в данном случае об атомах металлов
[13:37:04] Andrei: ну дак и ежу понятно что и другие атомы тоже могут иметь подобное строение
[13:38:30] Andrei: вернее не кольцегранную а многогранную
[13:38:51] Andrei: по сути это одно и то же
[13:40:06] Andrei: а про вращение спирали белка - это тоже не новость
[13:40:40] Andrei: так что новизна стремится к нулю
[13:44:02] Кушелев Александр Юрьевич: Я же говорю, что для Вас без разницы, кольцегранная, многогранная... молекула, кристалл... Пока не научитесь отличать молекулу от кристалла, я не готов с Вами дискутировать на научные темы...
[13:44:44] Кушелев Александр Юрьевич: Что касается "вращения спирали белка", то для Вас, похоже и ДНК от белка неотличима
[13:45:48] Andrei: ДНК это типичный пример вращения.. и ежу понятно что и в других белках тоже самое может происходить
[13:50:26] Кушелев Александр Юрьевич: Вы мне напомнили один армейский анекдот: "Копать будете от меня и до следующего дуба"
2017.01.24 21:26:12
>ENA|AAD33871|AAD33871.1 Homo sapiens (human) alpha-tubulin
ATGCGTGAGTGCATCTCCATCCACGTTGGCCAGGCTGGTGTCCAGATTGGCAATGCCTGC
TGGGAGCTCTACTGCCTGGAACACGGCATCCAGCCCGATGGCCAGATGCCAAGTGACAAG
ACCATTGGGGGAGGAGATGATTCCTTCAACACCTTCTTCAGTGAGACGGGGGCTGGCAAG
CATGTGCCCCGGGCAGTGTTTGTAGACTTGGAACCCACAGTCATTGATGAAGTTCGCACT
GGCACCTACCGCCAGCTCTTCCACCCTGAGCAACTTATCACAGGCAAAGAAGATGCTGCC
AATAACTATGCCCGAGGGCACTACACCATTGGCAAGGAGATCATTGACCTCGTGTTGGAC
CGAATTCGCAAGCTGGCCGACCAGTGCACGGGTCTCCAGGGCTTCTTGGTTTTCCACAGC
TTTGGTGGGGGAACTGGTTCTGGGTTCACCTCGCTGCTCATGGAACGTCTCTCAGTTGAT
TATGGCAAGAAGTCCAAGCTGGAGTTCTCTATTTACCCGGCGCCCCAGGTTTCCACAGCT
GTAGTTGAGCCCTACAACTCCATCCTCACCACCCACACCACCCTGGAGCACTCTGATTGT
GCCTTCATGGTAGACAATGAGGCCATCTATGACATCTGTCGTAGAAACCTCGATATTGAG
CGTCCAACCTATACTAACCTGAATAGGTTAATAGGTCAAATTGTGTCCTCCATCACTGCT
TCCCTGAGATTTGATGGAGCCCTGAATGTTGACCTGACAGAATTCCAGACCAACCTGGTG
CCCTATCCCCGCATCCACTTCCCTCTGGCCACATATGCCCCTGTCATCTCTGCTGAGAAA
GCCTACCATGAACAGCTTTCTGTAGCAGAGATCACCAATGCTTGCTTTGAGCCAGCCAAC
CAGATGGTGAAATGTGACCCTCGCCATGGTAAATACATGGCTTGCTGCCTGTTGTACCGT
GGTGACGTGGTTCCCAAAGATGTCAATGCTGCCATTGCCACCATCAAGACCAAGCGTACC
ATCCAGTTTGTGGATTGGTGCCCCACTGGCTTCAAGGTTGGCATCAACTACCAGCCTCCC
ACTGTGGTGCCTGGTGGAGACCTGGCCAAGGTACAGAGAGCTGTGTGCATGCTGAGCAAC
ACCACAGCCATTGCTGAGGCCTGGGCTCGCCTGGACCACAAGTTTGACCTGATGTATGCC
AAACGTGCCTTTGTTCACTGGTACGTTGGGGAGGGGATGGAGGAAGGTGAGTTTTCAGAG
GCCCGTGAGGACATGGCTGCCCTTGAGAAGGATTATGAGGAGGTTGGTGTGGATTCTGTT
GAAGGAGAGGGTGAGGAAGAAGGAGAGGAATAC
Композиционный код-4:
1,3,1,1,1,1,1,1,3,1,1,3,3,1,1,3,1,3,1,1,1,1,1,1,1,4,3,1,1,1,1,1,3,1,1,1,2,3,1,1,1,3,4,2,2,3,3,1,1,1,
1,1,1,3,1,4,4,3,1,1,3,4,1,4,2,4,3,2,1,1,3,1,2,1,3,3,3,3,1,3,1,1,1,1,1,1,1,1,3,1,3,3,1,2,1,3,3,3,3,1,
3,1,3,1,2,4,1,1,1,3,1,1,1,1,3,1,1,4,1,1,2,3,1,1,4,1,1,1,1,4,3,1,1,1,1,1,3,1,1,1,3,3,4,2,3,3,3,4,1,1,
4,4,1,1,3,3,1,2,3,3,3,1,1,1,1,1,4,1,1,3,3,1,4,4,1,1,3,1,2,3,2,3,1,1,1,1,1,1,1,1,1,1,1,1,4,1,1,3,3,3,
1,1,1,2,1,3,1,1,1,3,1,1,3,3,3,1,1,3,3,1,3,2,1,3,3,1,4,3,1,3,2,3,3,3,4,1,1,1,3,3,1,4,3,3,3,2,1,4,3,3,
1,4,2,3,1,1,1,1,4,4,1,3,1,1,1,1,1,3,4,1,2,3,1,3,1,1,3,3,1,3,1,1,3,3,1,3,3,2,2,1,1,1,3,3,1,3,1,2,1,1,
1,1,4,3,3,1,3,1,3,3,3,1,1,3,1,1,4,1,1,3,3,1,4,3,1,3,3,1,3,3,1,3,1,1,1,1,1,1,3,1,1,1,3,4,3,1,1,1,3,1,
1,1,3,1,1,1,1,1,3,1,3,4,4,3,3,2,1,4,1,1,2,1,3,3,4,1,1,4,1,1,1,2,1,3,3,1,1,1,3,1,4,1,1,1,3,1,4,1,3,1,
3,3,1,3,3,1,1,1,3,4,1,4,1,1,3,3,1,3,2,1,1,3,1,1,1,3,1,3,1,1,3,3,1,1,3,3,4,3,3,3,3,2,1,3,1,3,3,2,1,3,
Композиционный код-6:
5,3,1,1,1,1,1,1,3,5,5,3,3,5,5,3,5,3,1,1,1,1,1,1,1,4,3,1,1,1,1,1,3,5,5,5,2,3,5,5,5,3,5,2,2,3,3,1,1,1,
1,1,1,3,4,4,4,3,5,5,3,4,4,4,2,5,3,2,5,5,3,5,2,5,3,3,3,3,5,3,1,1,1,1,1,1,1,1,3,5,3,3,5,2,5,3,3,3,3,5,
3,5,3,5,2,4,1,1,1,3,1,1,1,1,3,1,1,4,1,1,2,3,1,1,4,1,1,1,1,4,3,1,1,1,1,1,3,5,5,5,3,3,5,2,3,3,3,4,1,1,
4,4,1,1,3,3,5,2,3,3,3,1,1,1,1,1,4,1,1,3,3,1,4,4,1,1,3,5,2,3,2,3,1,1,1,1,1,1,1,1,1,1,1,1,4,1,1,3,3,3,
5,5,5,2,5,3,5,5,5,3,5,5,3,3,3,5,5,3,3,5,3,2,5,3,3,5,5,3,5,3,2,3,3,3,4,1,1,1,3,3,5,5,3,3,3,2,5,5,3,3,
5,5,2,3,1,1,1,1,4,4,1,3,1,1,1,1,1,3,5,5,2,3,5,3,5,5,3,3,5,3,5,5,3,3,5,3,3,2,2,5,5,5,3,3,5,3,5,2,1,1,
1,6,4,3,3,5,3,5,3,3,3,5,5,3,1,1,4,1,1,3,3,5,5,3,5,3,3,5,3,3,5,3,1,1,1,1,1,1,3,5,5,5,3,5,3,5,5,5,3,5,
5,5,3,1,1,1,1,1,3,5,3,5,5,3,3,2,1,4,1,1,2,5,3,3,4,1,6,4,1,1,1,2,5,3,3,5,5,5,3,1,4,1,1,1,3,5,5,5,3,5,
3,3,5,3,3,5,5,5,3,4,1,4,6,1,3,3,5,3,2,5,5,3,5,5,5,3,5,3,5,5,3,3,5,5,3,3,5,3,3,3,3,2,5,3,5,3,3,2,5,3,
Вторичная структура (Пикотех-2017)

В структуре первой субъединицы встречается 3 метионина.
>ENA|AAD33873|AAD33873.1 Homo sapiens (human) beta-tubulin
ATGAGGGAAATCGTGCACATCCAGGCTGGTCAGTGTGGCAACCAGATCGGTGCCAAGTTC
TGGGAGGTGATCAGTGATGAACATGGCATCGACCCCACCGGCACCTACCACGGGGACAGC
GACCTGCAGCTGGACCGCATCTCTGTGTACTACAATGAAGCCACAGGTGGCAAATATGTT
CCTCGTGCCATCCTGGTGGATCTAGAACCTGGGACCATGGACTCTGTTCGCTCAGGTCCT
TTTGGCCAGATCTTTAGACCAGACAACTTTGTATTTGGTCAGTCTGGGGCAGGTAACAAC
TGGGCCAAAGGCCACTACACAGAGGGCGCCGAGCTGGTTGATTCTGTCCTGGATGTGGTA
CGGAAGGAGGCAGAGAGCTGTGACTGCCTGCAGGGCTTCCAGCTGACCCACTCACTGGGC
GGGGGCACAGGCTCTGGAATGGGCACTCTCCTTATCAGCAAGATCCGAGAAGAATACCCT
GATCGCATCATGAATACCTTCAGTGTGGTGCCTTCACCCAAAGTGTCTGACACCGTGGTC
GAGCCCTACAATGCCACCCTCTCCGTCCATCAGTTGGTAGAGAATACTGATGAGACCTAT
TGCATTGACAACGAGGCCCTCTATGATATCTGCTTCCGCACTCTGAAGCTGACCACACCA
ACCTACGGGGATCTGAACCACCTTGTCTCAGCCACCATGAGTGGTGTCACCACCTGCCTC
CGTTTCCCTGGCCAGCTCAATGCTGACCTCCGCAAGTTGGCAGTCAACATGGTCCCCTTC
CCACGTCTCCATTTCTTTATGCCTGGCTTTGCCCCTCTCACCAGCCGTGGAAGCCAGCAG
TATCGAGCTCTCACAGTGCCGGAACTCACCCAGCAGGTCTTCGATGCCAAGAACATGATG
GCTGCCTGTGACCCCCGCCACGGCCGATACCTCACCGTGGCTGCTGTCTTCCGTGGTCGG
ATGTCCATGAAGGAGGTCGATGAGCAGATGCTTAACGTGCAGAACAAGAACAGCAGCTAC
TTTGTGGAATGGATCCCCAACAATGTCAAGACAGCCGTCTGTGACATCCCACCTCGTGGC
CTCAAGATGGCAGTCACCTTCATTGGCAATAGCACAGCCATCCAGGAGCTCTTCAAGCGC
ATCTCGGAGCAGTTCACTGCCATGTTCCGCCGGAAGGCCTTCCTCCACTGGTACACAGGC
GAGGGCATGGACGAGATGGAGTTCACCGAGGCTGAGAGCAACATGAACGACCTCGTCTCT
GAGTATCAGCAGTACCAGGATGCCACCGCAGAAGAGGAGGAGGATTTCGGTGAGGAGGCC
GAAGAGGAGGCC
Композиционный код-4:
1,1,3,1,4,1,1,1,3,3,1,3,1,1,1,1,3,1,1,1,1,1,4,1,3,3,3,3,1,1,1,1,1,1,1,1,1,4,1,1,1,4,1,4,1,1,1,3,4,1,
1,3,3,1,2,3,1,3,3,3,3,3,1,1,4,4,3,2,3,3,4,1,1,1,3,3,1,2,3,3,3,1,1,1,3,3,2,1,1,3,2,3,3,1,3,4,2,3,1,1,
1,1,3,1,1,1,2,1,1,1,1,4,3,3,3,1,4,3,4,2,4,1,1,2,1,1,3,1,1,4,1,1,1,1,4,1,1,2,4,1,4,1,2,1,3,2,1,1,3,1,
3,1,1,1,1,2,3,3,1,3,3,1,1,1,3,1,1,3,4,4,3,2,1,3,4,3,1,1,4,1,1,1,1,3,1,1,1,1,1,3,1,1,2,1,3,3,3,1,1,3,
1,3,1,1,1,1,1,3,3,1,1,1,1,3,4,1,4,1,2,2,1,1,4,3,4,1,1,3,1,2,1,1,1,3,3,1,1,1,1,1,3,1,3,1,1,1,3,3,1,1,
1,1,1,2,1,1,1,1,1,1,2,3,1,3,1,3,1,3,1,3,1,3,1,1,1,3,2,1,1,1,3,2,3,1,2,4,4,3,1,1,1,1,1,1,3,1,1,1,1,1,
3,1,3,1,1,1,1,1,2,1,1,1,4,3,3,1,1,3,3,4,1,1,1,1,1,1,3,1,1,1,3,1,4,1,1,1,1,1,1,1,3,4,3,1,1,1,1,3,1,1,
2,1,1,3,1,1,2,3,3,1,1,1,1,2,1,1,1,3,1,3,1,2,1,1,1,1,1,1,1,1,1,4,1,1,1,3,1,1,1,1,4,1,1,1,1,1,1,1,2,1,
1,1,1,1,1,1,1,1,1,1,3,1,1,1,1,1,1,1,1,3,1,3,1,1,1,1,3,1,1,2,3,1,1,1,3,1,3,1,1,1,3,1,1,3,1,3,3,2,1,3,
Композиционный код-6:
5,5,3,1,4,1,1,1,3,3,5,3,1,1,1,1,3,1,1,1,1,1,4,1,3,3,3,3,1,1,1,1,1,1,1,1,1,4,1,1,1,4,1,4,1,1,1,3,5,5,
5,3,3,5,2,3,5,3,3,3,3,3,1,1,4,4,3,2,3,3,4,1,6,1,3,3,5,2,3,3,3,5,5,5,3,3,2,5,5,3,2,3,3,5,3,5,2,3,1,1,
1,1,3,5,5,5,2,1,1,1,1,4,3,3,3,5,5,3,5,2,5,5,5,2,5,5,3,1,1,4,1,1,1,1,4,1,1,2,4,1,4,1,2,5,3,2,5,5,3,5,
3,1,1,1,1,2,3,3,5,3,3,5,5,5,3,5,5,3,5,5,3,2,5,3,5,3,1,1,4,1,1,1,1,3,1,1,1,1,1,3,5,5,2,5,3,3,3,5,5,3,
5,3,1,1,1,1,1,3,3,1,1,1,1,3,4,1,4,1,2,2,4,4,4,3,5,5,5,3,5,2,5,5,5,3,3,1,1,1,1,1,3,5,3,5,5,5,3,3,1,1,
1,1,1,2,1,1,6,1,1,1,2,3,5,3,5,3,5,3,5,3,5,3,5,5,5,3,2,5,5,5,3,2,3,5,2,5,5,3,1,1,1,1,1,1,3,1,1,1,6,1,
3,5,3,1,1,1,1,1,2,1,1,1,4,3,3,5,5,3,3,4,6,1,6,1,1,1,3,5,5,5,3,1,4,1,1,1,1,1,1,1,3,5,3,1,1,1,1,3,5,5,
2,5,5,3,5,5,2,3,3,1,1,1,1,2,5,5,5,3,5,3,5,2,1,1,1,1,1,1,1,1,1,4,1,1,1,3,1,6,1,1,4,1,1,1,1,1,1,1,2,1,
1,1,6,1,1,6,1,1,1,1,3,1,1,1,6,1,1,1,1,3,5,3,1,1,1,1,3,5,5,2,3,5,5,5,3,5,3,5,5,5,3,5,5,3,5,3,3,2,5,3,

В структуре второй субъединицы встречается целых 9(девять!) Met (метионинов). На них спирали заметно гнутся. А на кодах "4" заметно (градусов на 20) скручиваются. Поэтому форма модели сильно отличается, если не учитывать эти изгибы и скручивания.
Victoriy пишет:
Мне для японцев нужен координатный файл гетеродимера, а использовать "сказочную" модель из базы данных не хочется. Моя программа виснет при работе с двумя белками сразу.
Ещё раз спрашиваю: в своих программах или вьюере Вы могли бы ориентировать две мои модели хотя бы рядом друг с другом. Мне туда ещё молекулы куркумина всунуть надо.
А дальше пусть они сами с этим играются.
Кушелев: Две модели можно крутить в программе HiperChem
Вы объясните заказчикам, что стуктуры этих белков содержат метионины и коды 310-спирали. Поэтому нужно учитывать скручивание и изгиб спиралей. Валентин делает версию Пикотех, которая поможет легко настроить средние углы и в интерактивном режиме поправлять модели конкретных белков вручную, т.е. в режиме пошаговой сборки модели.
А на модели из PDB, как мы с Вами понимаем, надеяться не приходится
2017.01.21 21:23:37
http://www.uniprot.org/uniprot/P09945

http://www.ebi.ac.uk/ena/data/view/AAA29195
>ENA|AAA29195|AAA29195.1 Hirudo medicinalis (medicinal leech) hypothetical protein
GCAATCTGCGTGTCTCAAGCAATTACTTACACTGATTGTACAGAATCGGGTCAAAATTTG
TGCCTCTGCGAGGGAAGCAATGTTTGCGGTAAAGGCAATAAGTGCATATTGGGTTCTAAT
GGAAAGGGCAACCAATGTGTCACTGGCGAAGGTACACCGAACCCTGAAAGCCATAATAAC
GGCGATTTCGAAGAAATTCCAGAAGAATATTTACAATGA

Композиционный код-6:
2,4,4,4,3,3,2,3,3,5,3,3,3,2,3,5,3,3,3,1,1,1,1,1,2,5,3,3,5,3,3,5,3,5,5,2,5,3,3,3,2,5,5,5,3,3,5,3,5,3,
3,2,5,5,3,3,5,3,3,5,5,3,5,3,3,3,2,3,3,3,3,3,0

Модель построена с помощью скрипта Кушелева-2017 по композиционному коду-6
Модель показывает, что цикл замыкается через Cys3...Cys21
2017.01.21 21:08:18
http://www.rcsb.org/pdb/explore/explore … ureId=1A4W
http://www.uniprot.org/uniprot/P00734
http://www.ebi.ac.uk/ena/data/view/CAJ01369
>ENA|CAJ01369|CAJ01369.1 Homo sapiens (human) prothrombin
ATGGCGCACGTCCGAGGCTTGCAGCTGCCTGGCTGCCTGGCCCTGGCTGCCCTGTGTAGC
CTTGTGCACAGCCAGCATGTGTTCCTGGCTCCTCAGCAAGCACGGTCGCTGCTCCAGCGG
GTCCGGCGAGCCAACACCTTCTTGGAGGAGGTGCGCAAGGGCAACCTAGAGCGAGAGTGC
GTGGAGGAGACGTGCAGCTACGAGGAGGCCTTCGGGGCTCTGGAGTCCTCCACGGCTACG
GATGTGTTCTGGGCCAAGTACACAGCTTGTGAGACAGCGAGGACGCCTCGAGATAAGCTT
GCTGCATGTCTGGAAGGTAACTGTGCTGAGGGTCTGGGTACGAACTACCGAGGGCATGTG
AACATCACCCGGTCAGGCATTGAGTGCCAGCTATGGAGGAGTCGCTACCCACATAAGCCT
GAAATCAACTCCACTACCCATCCTGGGGCCGACCTACAGGAGAATTTCTGCCGCAACCCC
GACAGCAGCACCACGGGACCCTGGTGCTACACTACAGACCCCACCGTGAGGAGGCAGGAA
TGCAGCATCCCTGTCTGTGGCCAGGATCAAGTCACTGTAGCGATGACTCCACGCTCCGAA
GGCTCCAGTGTGAATCTGTCACCTCCATTGGAGCAGTGTGTCCCTGATCGGGGGCAGCAG
TACCAGGGGCGCCTGGCGGTGACCACACATGGGCTCCCCTGCCTGGCCTGGGCCAGCGCA
CAGGCCAAGGCCCTGAGCAAGCACCAGGACTTCAACTCAGCTGTGCAGCTGGTGGAGAAC
TTCTGCCGCAACCCAGACGGGGATGAGGAGGGCGTGTGGTGCTATGTGGCCGGGAAGCCT
GGCGACTTTGGGTACTGCGACCTCAACTATTGTGAGGAGGCCGTGGAGGAGGAGACAGGA
GATGGGCTGGATGAGGACTCAGACAGGGCCATCGAAGGGCGTACCGCCACCAGTGAGTAC
CAGACTTTCTTCAATCCGAGGACCTTTGGCTCGGGAGAGGCAGACTGTGGGCTGCGACCT
CTGTTCGAGAAGAAGTCGCTGGAGGACAAAACCGAAAGAGAGCTCCTGGAATCCTACATC
GACGGGCGCATTGTGGAGGGCTCGGATGCAGAGATCGGCATGTCACCTTGGCAGGTGATG
CTTTTCCGGAAGAGTCCCCAGGAGCTGCTGTGTGGGGCCAGCCTCATCAGTGACCGCTGG
GTCCTCACCGCCGCCCACTGCCTCCTGTACCCGCCCTGGGACAAGAACTTCACCGAGAAT
GACCTTCTGGTGCGCATTGGCAAGCACTCCCGCACAAGGTACGAGCGAAACATTGAAAAG
ATATCCATGTTGGAAAAGATCTACATCCACCCCAGGTACAACTGGCGGGAGAACCTGGAC
CGGGACATTGCCCTGATGAAGCTGAAGAAGCCTGTTGCCTTCAGTGACTACATTCACCCT
GTGTGTCTGCCCGACAGGGAGACGGCAGCCAGCTTGCTCCAGGCTGGATACAAGGGGCGG
GTGACAGGCTGGGGCAACCTGAAGGAGACGTGGACAGCCAACGTTGGTAAGGGGCAGCCC
AGTGTCCTGCAGGTGGTGAACCTGCCCATTGTGGAGCGGCCGGTCTGCAAGGACTCCACC
CGGATCCGCATCACTGACAACATGTTCTGTGCTGGTTACAAGCCTGATGAAGGGAAACGA
GGGGATGCCTGTGAAGGTGACAGTGGGGGACCCTTTGTCATGAAGAGCCCCTTTAACAAC
CGCTGGTATCAAATGGGCATCGTCTCATGGGGTGAAGGCTGTGACCGGGATGGGAAATAT
GGCTTCTACACACATGTGTTCCGCCTGAAGAAGTGGATACAGAAGGTCATTGATCAGTTT
GGAGAGTAG

2017.01.22 21:17:08
[20:29:35] Кушелев Александр Юрьевич: Вот что я писал в 2012-ом: "Вот он, правильный инсулин.
Только теперь его удалось получить меняя не три, а всего лишь один композиционный угол!"
[20:29:48] Кушелев Александр Юрьевич: А на самом деле придётся делать по-настоящему...
[20:30:10] Va12220(valqwa): ну наконец то
[20:30:20] Va12220(valqwa): не зря я тебе мозги промывал ацетоном вчера
[20:31:20] Кушелев Александр Юрьевич:
[20:32:07] Кушелев Александр Юрьевич: Так меняется транспозиционный угол. Его придётся менять и для одного, и для всех аминокислот в реальном времени. Сейчас покажу, как ты такое делал в 2012-ом...
[20:32:46] Кушелев Александр Юрьевич:

[20:33:17] Кушелев Александр Юрьевич: Это я менял один угол альфа-спирали для всех аминокислотных остатков сразу
[20:33:59] Кушелев Александр Юрьевич: А надо будет сделать не только "1", но и "23456" и транспозиционный тоже. И тоже в реал тайм.
[20:34:13] Кушелев Александр Юрьевич: Но сначала нужно собрать алгоритм правильно.
[20:35:23] Кушелев Александр Юрьевич: Давай для начала поставим правильно первый и второй аминокислотные остатки.
[20:35:55] Кушелев Александр Юрьевич: И потом подумаем, как последний ставить на место предпоследнего в случае альфа-спирали.
[20:36:45] Кушелев Александр Юрьевич: А в случае не альфа-спирали нужно будет делать реверс последнего остатка, но уже вместе с построенной частью белка
[20:37:14] Кушелев Александр Юрьевич: А потом переводить как бы в положение предпоследнего, если бы это была альфа-спираль.
[20:39:56] Кушелев Александр Юрьевич: Короче, возвращаемся сюда и делаем в твоей программе такую модель:
[20:39:57] Кушелев Александр Юрьевич:

Тут координаты
[20:40:25] Va12220(valqwa): серые шарики что обозначают ?
[20:40:43] Кушелев Александр Юрьевич: Центры электронов-колец
[20:41:02] Кушелев Александр Юрьевич: А более крупные - ядра углерода
[20:41:14] Кушелев Александр Юрьевич: Синий - азот
[20:41:15] Va12220(valqwa): а голубой шарик без колец там у тебя
[20:41:54] Кушелев Александр Юрьевич: А голубой - кислород, который будет вытолкнут (замещён) азотом последней аминокислоты сверху вниз
[20:42:42] Кушелев Александр Юрьевич: Голубой шарик должен находиться в начале координат: (0,0,0)
[20:42:55] Кушелев Александр Юрьевич: Но вращаться должна сначала верхняя аминокислота
[20:43:03] Va12220(valqwa): но колечки вокруг него тоже нужны и важны
[20:43:08] Кушелев Александр Юрьевич: А после установки должен быть реверс всей структуры
[20:43:19] Кушелев Александр Юрьевич: Колечки будут
[20:43:27] Кушелев Александр Юрьевич: Ведь на это место встанет азот с колечками
[20:44:04] Кушелев Александр Юрьевич: Верхняя (последняя аминокислота) должна опускаться в таком положении:
[20:44:26] Кушелев Александр Юрьевич:

[20:45:13] Кушелев Александр Юрьевич: И перед тем, как её опустить, нужно её повернуть по варианту 123456, опустить
[20:46:10] Кушелев Александр Юрьевич: А потом объединить с нижней (и всем белком), сделать реверс в позицию 1 (альфа-спираль, начало отсчёта угла), после чего последний аминокислотный остаток вместе со всем белком поставить на место предпоследнего.
[20:46:43] Кушелев Александр Юрьевич: Это будет модель реального процесса в рибосоме.
[20:50:04] Кушелев Александр Юрьевич: Чтобы получить нулевой угол (угол альфа-спирали) нужно нижнюю модель перевести в положение верхней так, чтобы они были в композиции альфа-спирали
[20:59:44] Кушелев Александр Юрьевич: Верхний остаток нужно будет вместе с нижним потом повернуть так, чтобы ось иксов показывала нулевой угол для верхнего. А нижний будет ориентирован "как получится"
[21:01:42] Кушелев Александр Юрьевич: Угол альфа-спирали нужно будет настраивать параллельно с транспозиционным углом. Иначе он неправильным будет...
[21:03:10] Кушелев Александр Юрьевич: И после настройки сделать его нулевым для верхнего (последнего) аминокислотного остатка. Для этого нужно будет оба остатка синхронно повернуть так, чтобы ось иксов показала нулевой угол поворота для верхнего.
[21:03:36] Кушелев Александр Юрьевич: И тогда мы будем знать правильное положение нижнего.
2017.01.21 18:50:06

Замена элементов приводит к замене вида вторичной структуры.

2017.01.20 18:52:38
diprospan пишет:
моделирование третичной (пространственной) структуры белка всегда натыкается
на невозможность определения точной истинной конформации исследуемого белка.
ещё в 1968 году Сайрус Левинталь вычислил, что для белка, который имеет 100
аминокислотных остатков существует приблизительно 100 в сотой степени
конформаций, при этом за ничтожное время (10 в -13 степени секунды)
аминокислотная цепь формирует наиболее энергетически выгодный вариант.
https://ru.wikipedia.org/wiki/Парадокс_Левинталя
специалисты страшно мучаются с этим обстоятельством не в силах это постичь, и
моделируют туго-эмпирическим методом с привлечением слегка похожих гомологов
из разных перепутанных баз данных, но и все 3D структуры гомологов были некогда
выявлены не совсем точно эмпирически от фонаря, в итоге они получают мрак и хаос.
понимая всю обречённость своего существования специалисты тогда на худой конец
используют для перебора возможных конформаций специальные сервисы, которые
работают через суперкомпьютеры, предназначенные для выполнения подобных задач.
это тот случай когда белок умнее его изучающих специалистов, белок выбирает свою
нативную структуру (native state) среди бесчисленного множества возможных,
для цепи из 100 остатков их полный перебор занял бы 10^80 лет, если один переход
осуществлять за 10^−13 секунды, поэтому сложность проблемы заключается в том,
что данный вопрос нельзя решить экспериментально, так как придётся ждать 10^80 лет.
специалистам остаётся успокаивать себя тем, что белки могут сворачиваться разными
путями, "не обязательно следуя глобально оптимальному пути".
да и в самом деле, омлет отличается от яичницы глазуньи. )
не в силах постичь третичную структуру, специалисты смело переходят к четвертичной.
https://ru.wikipedia.org/wiki/Четвертичная_структура
а там и до пятеричной видимо не очень далеко.)
Спасибо Diprospan, меня тоже занимал этот парадокс, однако стоит изменить постановку задачи и никакого парадокса нет.
Смотрите: сейчас все не обосновано исходят из того, что сворачивание каждой молекулы новосинтезированного белка происходит из вытянутого плоского шнурка с бусиками аминокислот (недоразумение №1), поскольку пространственная структура не кодируется (недоразумение №2).
А теперь давайте будем исходить из того, что 3D-структура белка закодирована в геноме, поэтому она транслируется рибосомой также как первичная структура белка. В результате рибосома штампует для всех молекул одного белка абсолютно одинаковые структурные шаблоны с уникальной специфичной только для этого белка пространственной структурой. В процессе фолдинга остаётся только немного довести эту структуру до совершенства в соответствии с микроокружением (например спрятать все гидрофобные группы внутрь белковой глобулы). Согласитесь, что если задана уникальная структура отсчета (декодированный структурный шаблон), то из неё получить функциональную конформацию пара пустяков. Очевидно, что для всех молекул другого белка синтезируется именно их структурные шаблоны, у которых также короткий и однозначный путь к функциональной конформации. У белков в клетке это очень быстрый и активный процесс, поскольку, если не успел вовремя свернуться, тебя тут же признают бракованным и разберут на зап.части. В действительности 60-70 % новосинтезированных белков тут же разрушаются протеазами: сами понимаете какая огромная конкуренция остаться в живых.
2017.01.20 21:04:05
Victoriy пишет:
зачем Вам понадобились БАКТЕРИАЛЬНЫЕ белки. Зайдите в Гугл и узнайте, что бактериальный геном транслируется отлично от животных и растений: у них своя упрощенная система генетического кода даже первичной структуры белков, о пространственной я вообще молчу. Ни Вы, ни я не занимались вопросом особенностей кодирования пространственной структуры бактериальных белков. У них даже тРНК другие. Переносить на их транслирующую систему наработки, сделанные для эукариот -- бОльшая ошибка и небрежность с Вашей стороны.
Кушелев: А человеческие белки Вас устраивают?

2017.01.20 21:11:36
Вот ещё несколько человеческих, не гипотетических:
https://www.ncbi.nlm.nih.gov/protein/10 … 4MPRE7J015
https://www.ncbi.nlm.nih.gov/protein/55 … 4MPRE7J015
https://www.ncbi.nlm.nih.gov/protein/10 … 4MPRE7J015
https://www.ncbi.nlm.nih.gov/protein/78 … 4MPRE7J015
https://www.ncbi.nlm.nih.gov/protein/10 … 4MPRE7J015
https://www.ncbi.nlm.nih.gov/protein/45 … 4MPRE7J015
https://www.ncbi.nlm.nih.gov/protein/96 … 4MPRE7J015
https://www.ncbi.nlm.nih.gov/protein/45 … 4MPRE7J015
https://www.ncbi.nlm.nih.gov/nuccore/427197606
Вторичная структура (Пикотех-2017): https://cloud.mail.ru/public/EW1F/6rDqp29Ei

Два витка метиониновой спирали
2017.01.20 22:05:03
А этот белок содержит длинную аланиновую спираль:
https://www.ncbi.nlm.nih.gov/protein/15 … 4MPRE7J015
2017.01.20 22:11:27
Kushelev пишет:
Вот ещё несколько человеческих, не гипотетических:
https://www.ncbi.nlm.nih.gov/protein/10 … 4MPRE7J015
https://www.ncbi.nlm.nih.gov/protein/55 … 4MPRE7J015
https://www.ncbi.nlm.nih.gov/protein/45 … 4MPRE7J015
https://www.ncbi.nlm.nih.gov/nuccore/427197606
Вторичная структура (Пикотех-2017): https://cloud.mail.ru/public/EW1F/6rDqp29Ei
А.Ю., некоторое повторение метионинов на N-конце приведенных Вами белков могло случиться в ходе их эволюции посредством дупликации кодона сигнального N-концевого метионина любого белка, который как правило отщепляется в ходе его посттрансляционного созревания.
Действительно в приведенных Вами примерах есть не один метионин в виде затравки трансляции, а 4-7 штук. Подозреваю, что такая множественная дупликация не мешала ни их синтезу, ни структуре, поскольку метиониновый хвост НЕ сворачивался в виток спирали, как об этом свидетельствуют экспериментальные данные и результаты моей программы: http://www.uniprot.org/uniprot/Q12824.
В белке http://www.uniprot.org/uniprot/Q01668 полиметионин (1-7), полилейцин (653-659) и полиглутамат (827-838) используются не в качестве мифических спиралей Кушелева, а как связующие мостики для композиционного смещения топологических и трансмембранных доменов этого огромного белка.
Victoriy пишет:
Действительно в приведенных Вами примерах есть не один метионин в виде затравки трансляции, а 4-7 штук. Подозреваю, что такая множественная дупликация не мешала ни их синтезу, ни структуре, поскольку метиониновый хвост НЕ сворачивался в виток спирали, как об этом свидетельствуют экспериментальные данные
Кушелев: А где можно посмотреть на эти экспериментальные данные?
И как быть с метиониновой спиралью в конце или в середине белковой молекулы?
Это - не начало белка!
И это, как видите, далеко не начало белка
Если экспериментальные данные относятся к синтетическим полипептидам, то композиционный код тут ни при чём. Другое дело, если метиониновые спирали синтезируются рибосомой. Пусть даже по синтетическому гену. Такие экспериментальные данные есть?
Кстати. никакая аминокислота, многократно повторенная в полимере, например, в бета-спирали, не будет сворачиваться в альфа-спираль. Построить альфа-спираль может только рибосома.
http://www.uniprot.org/uniprot/Q12824
http://www.ebi.ac.uk/ena/data/view/AAA81905
Вторичная структура (Пикотех-2017): https://img-fotki.yandex.ru/get/42692/1 … 1_orig.png
{kind=link}

С какими экспериментальными данными Вы предлагаете сравнить вторичную структуру по алгоритму Пикотех-2017 ?
С этими?
http://www.ebi.ac.uk/pdbe/entry/pdb/5AJ1

РСА "не видит" не только витка метиониновой спирали. Он вообще всю спираль от 1 до 32-го аминокислотного остатка "не видит" ![]()
Так что сравнивать-то не с чем. Нет правильных экспериментальных данных по участку 1..32 данного белка. РСА часто "не видит" до 50 аминокислотных остатков...
2017.01.20 22:52:01
Пикотехнология белков, ДНК, РНК
Victoriy пишет:
полиметионин (1-7), полилейцин (653-659) и полиглутамат (827-838) используются не в качестве мифических спиралей Кушелева, а как связующие мостики для композиционного смещения топологических и трансмембранных доменов этого огромного белка.
А что, по Вашей программе полилейцин и полиглутоамат тоже не могут иметь спиральную структуру?
http://www.uniprot.org/uniprot/Q01668
http://www.ebi.ac.uk/pdbe/entry/pdb/3LV3

http://www.rcsb.org/pdb/explore/explore … ureId=3LV3


Кушелев: Уважаемая Виктория! Если Вы этими "экспериментальными" данными оперируете, то они действительно не подтверждают существования метиониновой спирали. Однако они не подтверждают и другие альфа-спирали. Всё, что показано здесь красным цветом, эти "экспериментальные" данные не подтверждают. Верите? Но это не экспериментальные данные ![]()
http://www.ebi.ac.uk/ena/data/view/AAA58402
Вторичная структура (Пикотех-2017): https://cloud.mail.ru/public/3QW1/9ho83Y57A

2017.01.20 23:44:32
Пикотехнология белков, ДНК, РНК
Victoriy пишет:
В белке http://www.uniprot.org/uniprot/Q01668 полиметионин (1-7), полилейцин (653-659) и полиглутамат (827-838) используются не в качестве мифических спиралей Кушелева, а как связующие мостики для композиционного смещения топологических и трансмембранных доменов этого огромного белка.

Кушелев: По поводу полилейцина

и полиглутамата я и не утверждал, что это альфа-спирали
А метиониновая спираль - это спираль однозначно.
Кстати, уважаемая Виктория!
Вы заметили, что построены модели 6 замкнутых через дисульфидные мостики участков лизоцимов из разных организмов? И по Вашей просьбе я нашёл экспериментальные подтверждения...
1. Cys 95 - Cys 113 в человеческом лизоциме.
2. Cys 96 - Cys 117 в альтернативном человеческом лизоциме.
3. Cys 78 - Cys 82 в альтернативном человеческом лизоциме.
4. Cys 94 - Cys 98 в лизоциме куриного яйца
5. Met 1 - Cys 13 в мышином лизоциме
6. Met 1 - Cys 24 в лизоциме куриного яйца