2017.01.12 20:10:56
Формы, механизмы, энергия наномира
Пикотехнология белков, ДНК, РНК
http://www.rcsb.org/pdb/explore/explore … ureId=2ND1
http://www.uniprot.org/uniprot/O60885
http://www.ebi.ac.uk/ena/data/view/AAL26987
>ENA|AAL26987|AAL26987.1 Homo sapiens (human) bromodomain-containing 4
ATGTCTGCGGAGAGCGGCCCTGGGACGAGATTGAGAAATCTGCCAGTAATGGGGGATGGA
CTAGAAACTTCCCAAATGTCTACAACACAGGCCCAGGCCCAACCCCAGCCAGCCAACGCA
GCCAGCACCAACCCCCCGCCCCCAGAGACCTCCAACCCTAACAAGCCCAAGAGGCAGACC
AACCAACTGCAATACCTGCTCAGAGTGGTGCTCAAGACACTATGGAAACACCAGTTTGCA
TGGCCTTTCCAGCAGCCTGTGGATGCCGTCAAGCTGAACCTCCCTGATTACTATAAGATC
ATTAAAACGCCTATGGATATGGGAACAATAAAGAAGCGCTTGGAAAACAACTATTACTGG
AATGCTCAGGAATGTATCCAGGACTTCAACACTATGTTTACAAATTGTTACATCTACAAC
AAGCCTGGAGATGACATAGTCTTAATGGCAGAAGCTCTGGAAAAGCTCTTCTTGCAAAAA
ATAAATGAGCTACCCACAGAAGAAACCGAGATCATGATAGTCCAGGCAAAAGGAAGAGGA
CGTGGGAGGAAAGAAACAGGGACAGCAAAACCTGGCGTTTCCACGGTACCAAACACAACT
CAAGCATCGACTCCTCCGCAGACCCAGACCCCTCAGCCGAATCCTCCTCCTGTGCAGGCC
ACGCCTCACCCCTTCCCTGCCGTCACCCCGGACCTCATCGTCCAGACCCCTGTCATGACA
GTGGTGCCTCCCCAGCCACTGCAGACGCCCCCGCCAGTGCCCCCCCAGCCACAACCCCCA
CCCGCTCCAGCTCCCCAGCCCGTACAGAGCCACCCACCCATCATCGCGGCCACCCCACAG
CCTGTGAAGACAAAGAAGGGAGTGAAGAGGAAAGCAGACACCACCACCCCCACCACCATT
GACCCCATTCACGAGCCACCCTCGCTGCCCCCGGAGCCCAAGACCACCAAGCTGGGCCAG
CGGCGGGAGAGCAGCCGGCCTGTGAAACCTCCAAAGAAGGACGTGCCCGACTCTCAGCAG
CACCCAGCACCAGAGAAGAGCAGCAAGGTCTCGGAGCAGCTCAAGTGCTGCAGCGGCATC
CTCAAGGAGATGTTTGCCAAGAAGCACGCCGCCTACGCCTGGCCCTTCTACAAGCCTGTG
GACGTGGAGGCACTGGGCCTACACGACTACTGTGACATCATCAAGCACCCCATGGACATG
AGCACAATCAAGTCTAAACTGGAGGCCCGTGAGTACCGTGATGCTCAGGAGTTTGGTGCT
GACGTCCGATTGATGTTCTCCAACTGCTATAAGTACAACCCTCCTGACCATGAGGTGGTG
GCCATGGCCCGCAAGCTCCAGGATGTGTTCGAAATGCGCTTTGCCAAGATGCCGGACGAG
CCTGAGGAGCCAGTGGTGGCCGTGTCCTCCCCGGCAGTGCCCCCTCCCACCAAGGTTGTG
GCCCCGCCCTCATCCAGCGACAGCAGCAGCGATAGCTCCTCGGACAGTGACAGTTCGACT
GATGACTCTGAGGAGGAGCGAGCCCAGCGGCTGGCTGAGCTCCAGGAGCAGCTCAAAGCC
GTGCACGAGCAGCTTGCAGCCCTCTCTCAGCCCCAGCAGAACAAACCAAAGAAAAAGGAG
AAAGACAAGAAGGAAAAGAAAAAAGAAAAGCACAAAAGAAAAGAGGAAGTGGAAGAGAAT
AAAAAAAGCAAAGCCAAGGAACCTCCTCCTAAAAAGACGAAGAAAAATAATAGCAGCAAC
AGCAATGTGAGCAAGAAGGAGCCAGCGCCCATGAAGAGCAAGCCCCCTCCCACGTATGAG
TCGGAGGAAGAGGACAAGTGCAAGCCTATGTCCTATGAGGAGAAGCGGCAGCTCAGCTTG
GACATCAACAAGCTCCCCGGCGAGAAGCTGGGCCGCGTGGTGCACATCATCCAGTCACGG
GAGCCCTCCCTGAAGAATTCCAACCCCGACGAGATTGAAATCGACTTTGAGACCCTGAAG
CCGTCCACACTGCGTGAGCTGGAGCGCTATGTCACCTCCTGTTTGCGGAAGAAAAGGAAA
CCTCAAGCTGAGAAAGTTGATGTGATTGCCGGCTCCTCCAAGATGAAGGGCTTCTCGTCC
TCAGAGTCGGAGAGCTCCAGTGAGTCCAGCTCCTCTGACAGCGAAGACTCCGAAACAGAG
ATGGCTCCGAAGTCAAAAAAGAAGGGGCACCCCGGGAGGGAGCAGAAGAAGCACCATCAT
CACCACCATCAGCAGATGCAGCAGGCCCCGGCTCCTGTGCCCCAGCAGCCGCCCCCGCCT
CCCCAGCAGCCCCCACCGCCTCCACCTCCGCAGCAGCAACAGCAGCCGCCACCCCCGCCT
CCCCCACCCTCCATGCCGCAGCAGGCAGCCCCGGCGATGAAGTCCTCGCCCCCACCCTTC
ATTGCCACCCAGGTGCCCGTCCTGGAGCCCCAGCTCCCAGGCAGCGTCTTTGACCCCATC
GGCCACTTCACCCAGCCCATCCTGCACCTGCCGCAGCCTGAGCTGCCCCCTCACCTGCCC
CAGCCGCCTGAGCACAGCACTCCACCCCATCTCAACCAGCACGCAGTGGTCTCTCCTCCA
GCTTTGCACAACGCACTACCCCAGCAGCCATCACGGCCCAGCAACCGAGCCGCTGCCCTG
CCTCCCAAGCCCGCCCGGCCCCCAGCCGTGTCACCAGCCTTGACCCAAACACCCCTGCTC
CCACAGCCCCCCATGGCCCAACCCCCCCAAGTGCTGCTGGAGGATGAAGAGCCACCTGCC
CCACCCCTCACCTCCATGCAGATGCAGCTGTACCTGCAGCAGCTGCAGAAGGTGCAGCCC
CCTACGCCGCTACTCCCTTCCGTGAAGGTGCAGTCCCAGCCCCCACCCCCCCTGCCGCCC
CCACCCCACCCCTCTGTGCAGCAGCAGCTGCAGCAGCAGCCGCCACCACCCCCACCACCC
CAGCCCCAGCCTCCACCCCAGCAGCAGCATCAGCCCCCTCCACGGCCCGTGCACTTGCAG
CCCATGCAGTTTTCCACCCACATCCAACAGCCCCCGCCACCCCAGGGCCAGCAGCCCCCC
CATCCGCCCCCAGGCCAGCAGCCACCCCCGCCGCAGCCTGCCAAGCCTCAGCAAGTCATC
CAGCACCACCATTCACCCCGGCACCACAAGTCGGACCCCTACTCAACCGGTCACCTCCGC
GAAGCCCCCTCCCCGCTTATGATACATTCCCCCCAGATGTCACAGTTCCAGAGCCTGACC
CACCAGTCTCCACCCCAGCAAAACGTCCAGCCTAAGAAACAGGAGCTGCGTGCTGCCTCC
GTGGTCCAGCCCCAGCCCCTCGTGGTGGTGAAGGAGGAGAAGATCCACTCACCCATCATC
CGCAGCGAGCCCTTCAGCCCCTCGCTGCGGCCGGAGCCCCCCAAGCACCCGGAGAGCATC
AAGGCCCCCGTCCACCTGCCCCAGCGGCCGGAAATGAAGCCTGTGGATGTCGGGAGGCCT
GTGATCCGGCCCCCAGAGCAGAACGCACCGCCACCAGGGGCCCCTGACAAGGACAAACAG
AAACAGGAGCCGAAGACTCCAGTTGCGCCCAAAAAGGACCTGAAAATCAAGAACATGGGC
TCCTGGGCCAGCCTAGTGCAGAAGCATCCGACCACCCCCTCCTCCACAGCCAAGTCATCC
AGCGACAGCTTCGAGCAGTTCCGCCGCGCCGCTCGGGAGAAAGAGGAGCGTGAGAAGGCC
CTGAAGGCTCAGGCCGAGCACGCTGAGAAGGAGAAGGAGCGGCTGCGGCAGGAGCGCATG
AGGAGCCGAGAGGACGAGGATGCGCTGGAGCAGGCCCGGCGGGCCCATGAGGAGGCACGT
CGGCGCCAGGAGCAGCAGCAGCAGCAGCGCCAGGAGCAACAGCAGCAGCAGCAACAGCAA
GCAGCTGCGGTGGCTGCCGCCGCCACCCCACAGGCCCAGAGCTCCCAGCCCCAGTCCATG
CTGGACCAGCAGAGGGAGTTGGCCCGGAAGCGGGAGCAGGAGCGAAGACGCCGGGAAGCC
ATGGCAGCTACCATTGACATGAATTTCCAGAGTGATCTATTGTCAATATTTGAAGAAAAT
CTTTTCTGA
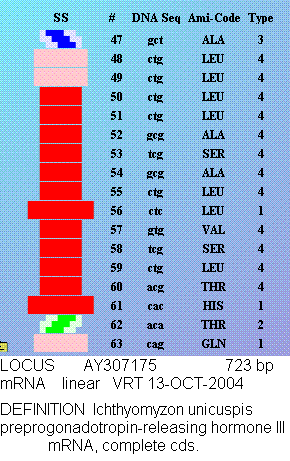
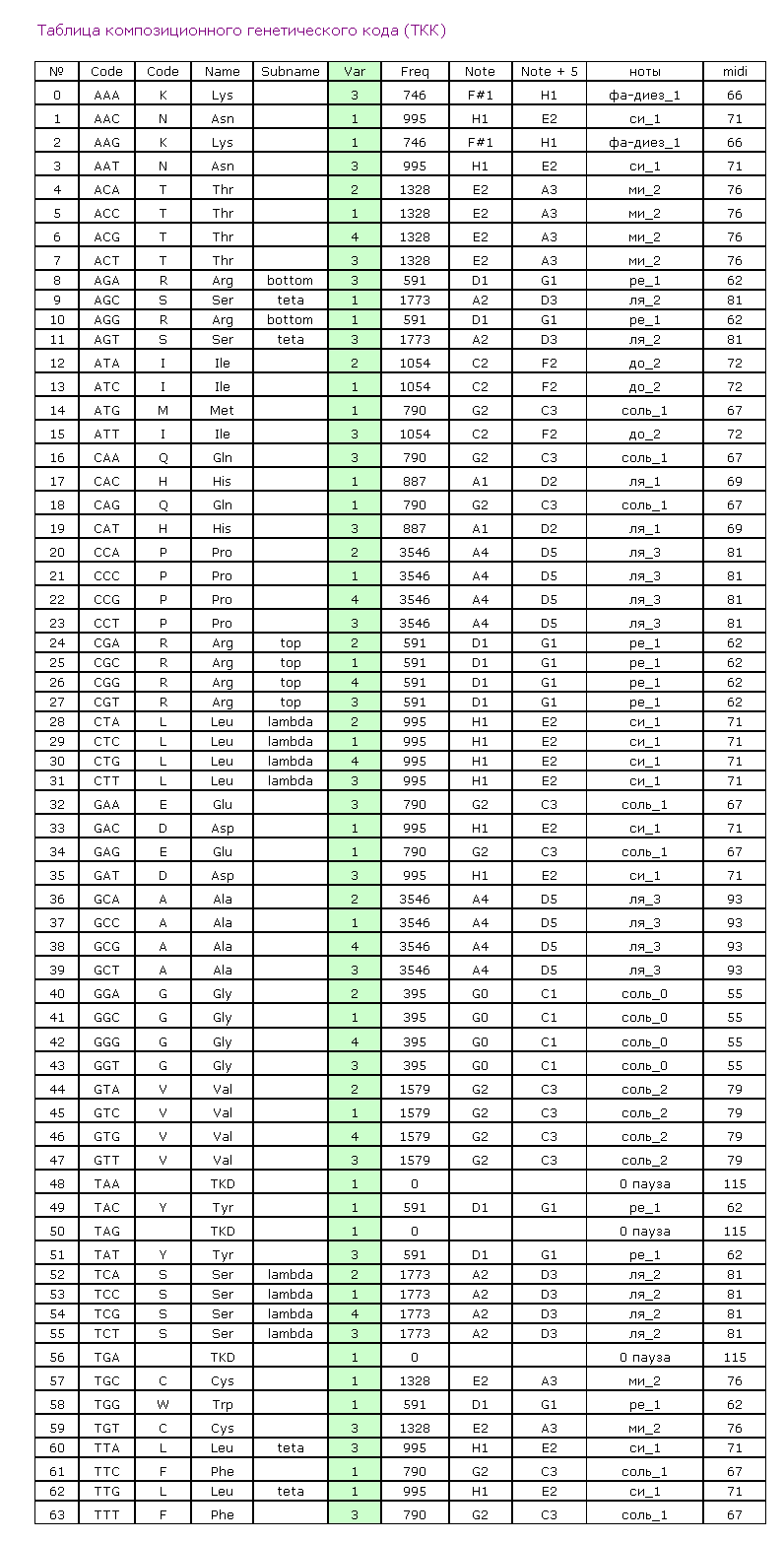
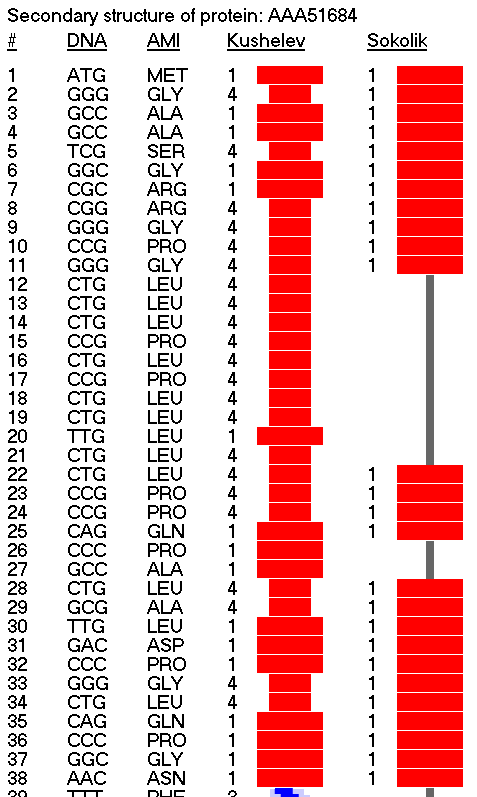
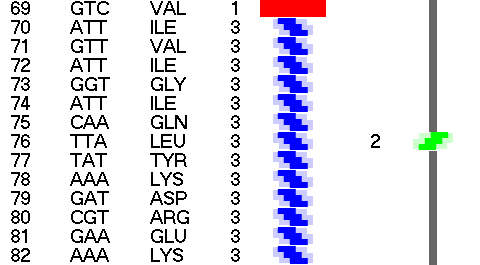
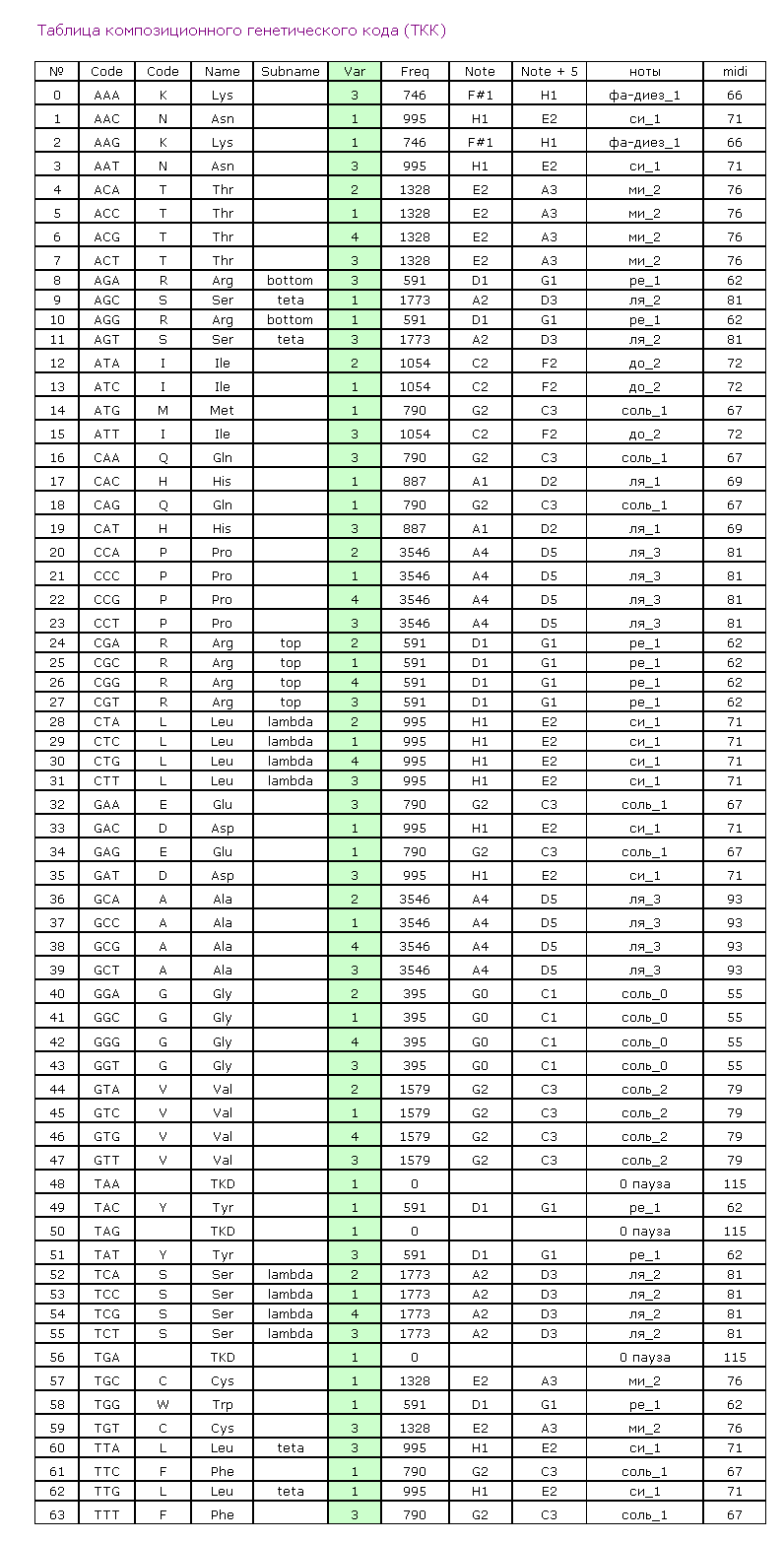
Композиционный код:
1,3,4,1,1,1,3,4,4,3,1,3,3,4,2,2,1,4,3,2,2,3,3,1,3,1,3,2,2,1,1,1,1,3,1,1,2,1,1,2,1,1,1,1,1,4,1,2,1,1,
1,1,3,1,1,1,1,1,1,1,1,3,4,3,1,4,1,3,4,4,1,1,2,2,1,3,1,1,3,2,1,3,1,1,1,3,4,3,1,1,1,4,1,1,3,3,1,3,1,1,
3,3,4,3,1,3,1,2,2,2,1,1,1,1,3,1,1,3,1,1,3,3,1,3,3,1,1,1,1,1,3,1,3,2,3,3,1,1,1,1,1,3,2,3,1,2,1,3,1,2,
3,3,4,3,1,1,1,1,3,3,2,3,1,2,1,2,3,3,1,1,1,1,2,1,1,2,3,2,3,2,3,4,1,3,3,2,4,2,2,3,3,1,3,1,4,2,2,1,2,3,
3,2,4,3,3,4,1,1,1,1,3,1,4,3,3,3,3,4,1,1,4,3,1,1,1,3,1,1,1,4,1,1,1,1,1,1,3,1,1,2,4,4,3,1,1,2,4,1,4,1,
4,2,4,1,1,1,2,3,1,2,1,3,2,3,1,1,1,2,1,1,1,2,1,1,1,4,1,1,2,1,3,4,1,2,1,1,2,4,1,1,3,2,1,1,1,1,1,1,1,3,
1,1,3,1,1,2,1,4,4,1,4,1,1,1,1,1,1,4,1,1,4,4,1,1,1,4,3,4,3,3,2,1,1,1,4,1,1,3,1,1,1,2,2,2,1,1,1,1,1,1,
4,1,1,1,1,1,1,1,1,1,1,1,1,1,3,1,1,1,1,1,1,1,1,1,1,1,1,1,3,4,1,4,1,2,4,1,2,1,1,1,3,1,1,1,1,1,1,1,1,1,
1,2,1,1,3,3,4,1,1,3,1,1,3,3,3,1,1,3,3,3,1,1,2,1,1,1,1,1,1,3,1,1,1,3,3,1,3,1,4,4,1,1,1,1,1,1,1,3,4,1,
3,1,1,3,1,1,1,4,1,1,3,1,1,2,4,4,1,4,1,1,4,2,4,1,3,1,1,1,3,4,1,4,1,2,1,1,1,1,1,1,3,1,1,4,1,3,1,3,4,3,
3,1,3,1,1,1,2,1,1,4,4,3,1,1,1,1,1,1,3,1,4,1,1,1,3,2,1,1,3,1,1,1,1,1,3,2,1,3,1,1,3,1,1,1,3,1,3,3,3,1,
1,3,3,3,1,3,4,3,1,3,3,3,1,3,1,1,3,3,3,3,3,1,4,1,3,3,3,1,1,1,1,3,4,1,1,1,1,2,4,1,1,1,1,1,1,3,1,4,3,1,
4,1,3,1,1,1,1,1,3,1,1,3,1,1,1,4,1,1,1,1,1,1,1,1,1,1,1,1,1,4,1,1,4,4,1,1,1,1,2,4,1,1,1,4,1,3,1,1,1,1,
1,3,3,1,1,3,1,1,4,1,4,1,2,4,3,1,4,1,1,3,1,1,1,3,1,4,1,3,1,3,3,3,3,1,3,3,3,4,3,1,1,1,1,1,1,1,1,1,4,1,
2,1,4,1,1,1,3,1,1,1,1,3,1,1,3,1,1,3,2,1,1,3,4,1,2,3,1,1,4,1,1,4,1,1,1,1,1,1,3,3,1,1,3,1,1,1,1,1,1,4,
3,3,4,1,1,1,4,1,4,3,1,1,1,1,2,4,3,2,3,4,1,1,3,1,1,4,2,1,4,3,1,2,1,1,1,4,1,1,2,1,4,4,1,1,1,4,1,2,1,1,
3,1,1,1,4,1,1,4,1,1,1,1,2,1,1,1,3,1,1,1,1,1,1,1,1,1,1,4,1,4,4,1,3,1,4,1,3,1,4,1,1,4,3,1,1,1,3,2,1,3,
1,1,1,1,2,4,1,3,3,2,3,1,1,1,2,2,1,1,1,2,2,4,1,1,1,2,1,3,1,4,3,1,1,1,1,4,1,2,1,4,2,2,1,1,1,3,2,1,4,1,
2,1,1,1,1,1,3,1,1,3,4,4,4,1,3,3,1,2,3,1,2,1,1,1,1,1,1,1,1,4,1,4,1,1,4,1,1,4,1,1,3,4,4,2,1,3,1,4,1,4,
1,1,1,1,2,1,1,4,4,1,2,1,1,1,3,4,1,1,1,4,1,1,1,4,2,2,1,2,2,1,1,1,1,3,2,1,1,1,1,3,1,1,3,2,4,1,4,1,1,1,
1,1,1,3,1,1,1,1,3,1,1,4,2,1,1,1,1,1,1,1,3,4,1,2,1,1,1,2,1,4,4,1,3,1,1,3,1,3,1,1,1,1,1,3,2,1,4,1,1,1,
4,1,1,1,2,1,3,1,1,1,3,1,1,1,4,3,1,2,3,1,1,1,1,2,1,1,1,1,4,1,1,1,3,2,1,1,3,1,1,1,3,1,3,1,1,4,3,3,1,1,
4,1,1,1,1,1,1,4,4,4,1,1,1,1,1,1,2,1,1,1,1,1,1,1,1,1,1,4,4,4,4,1,1,1,1,1,4,1,1,1,1,1,1,1,1,4,1,1,4,4,
3,1,1,3,4,3,1,4,1,3,4,1,4,1,2,1,1,1,2,4,2,2,4,1,3,1,1,1,3,1,3,1,1,4,1,3,2,3,4,1,3,1,1,4,3,1,1,1,1,1,
1,1,1,1,2,4,1,1,3,4,1,1,1,1,1,2,1,1,2,1,1,1,1,1,1,1,1,1,1,1,3,4,1,3,1,1,3,1,1,1,4,1,3,1,1,1,1,3,1,1,
1,1,1,4,4,4,1,1,1,1,1,1,2,1,1,1,3,4,4,1,1,1,4,4,1,3,1,1,2,3,4,1,1,1,1,1,1,1,1,1,1,1,3,1,1,1,1,3,1,3,
2,3,4,4,3,1,1,1,1,2,1,1,1,1,1,1,1,1,1,1,4,1,1,1,1,1,1,1,4,1,4,1,1,1,2,3,1,4,3,1,1,2,3,1,3,1,1,3,1,1,
3,3,2,1,2,2,3,3,3,3,3,1
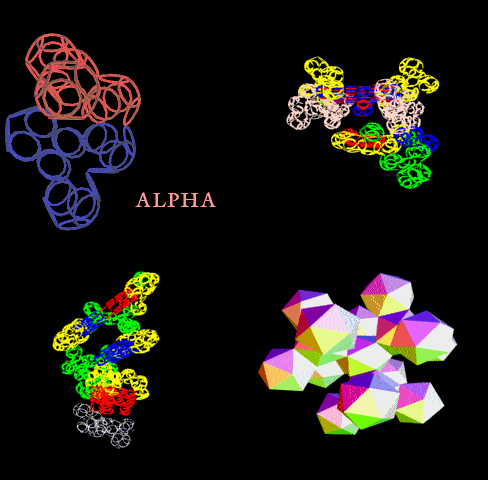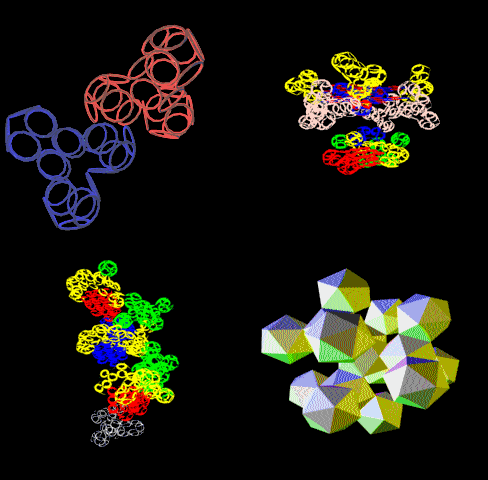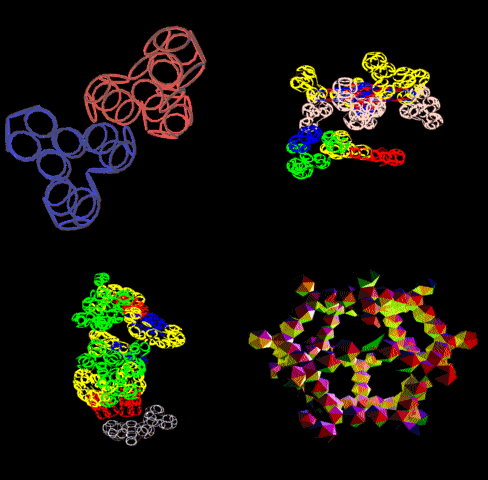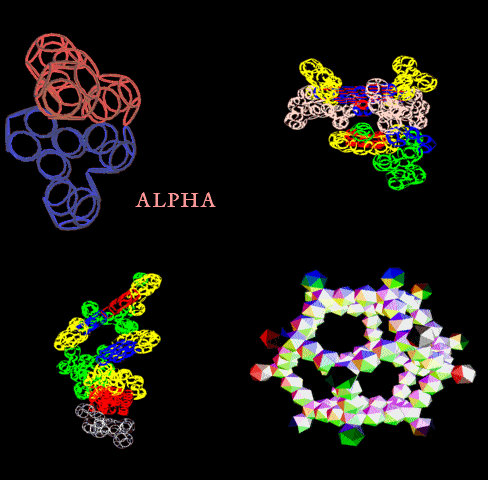
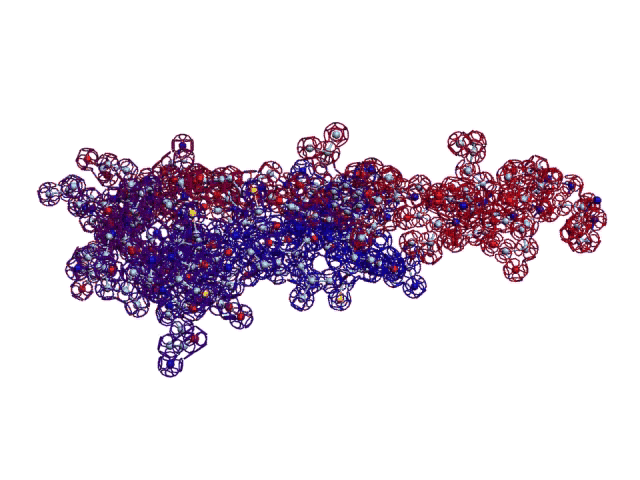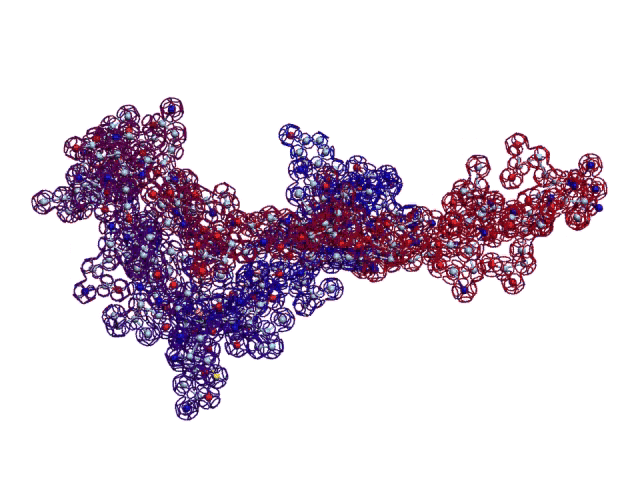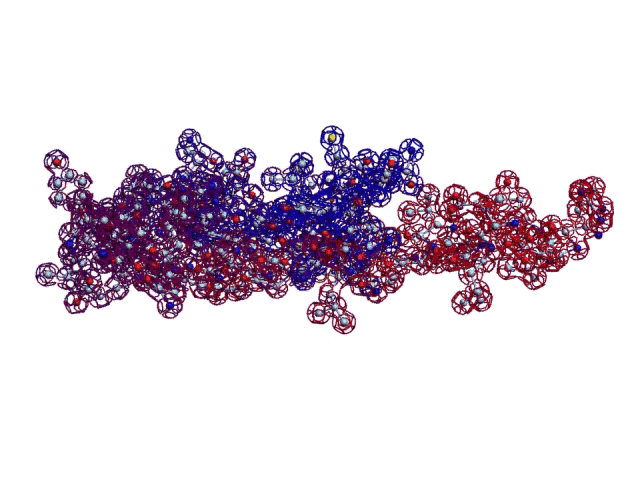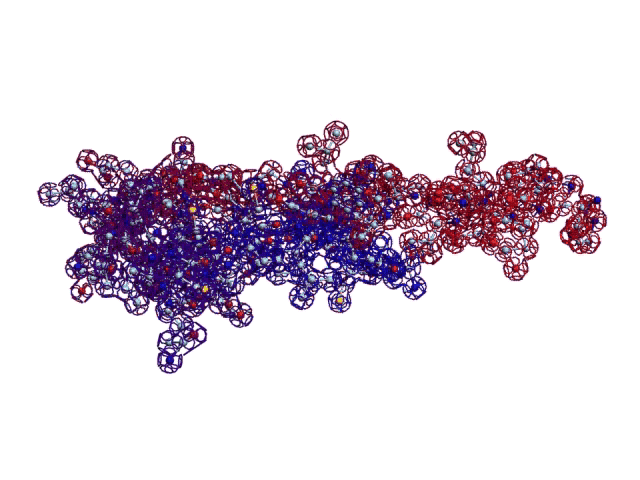
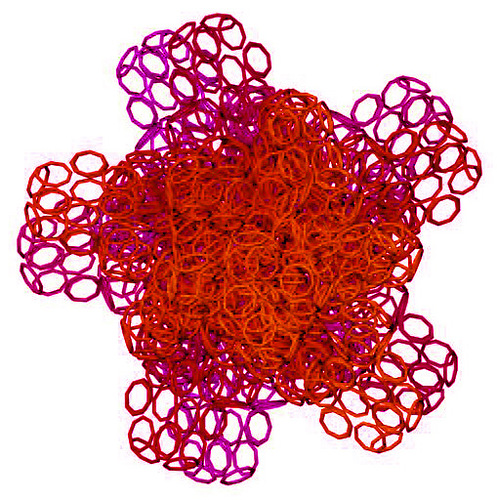
Вторичная структура (Пикотех): https://img-fotki.yandex.ru/get/195419/ … 4_orig.gif

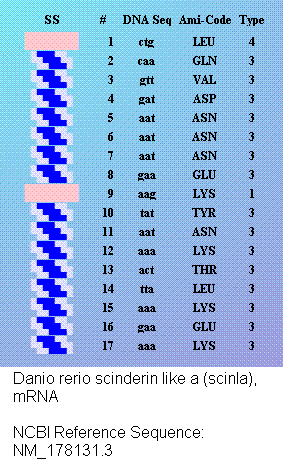
Кушелев: Любопытный фрагмент структуры...





Кушелев: Интересно, что это за участки белка такие необычные?

Кушелев: Обратите внимание на корреляцию вторичной структуры с аминокислотными остатками. Gln (глутамин) находится в структуре прямой альфа-спирали, а Pro (пролин) в структуре бета-спирали и краёв.
2017.01.12 20:41:06
Формы, механизмы, энергия наномира.
Пикотехнология белков, ДНК, РНК
Кушелев: Все видят корреляцию?

Такая корреляция наглядно показывает предсказательную силу пикотехнологии даже на уровне 2D-структуры.





Кушелев: Корреляция вторичной структуры по таблице композиционного кода с первичной последовательностью белка, в частности, иминокислоты Pro, очевидна.
А первичная последовательность в отличие от данных РСА имеет 100%-ную надёжность

































{kind=link}
{kind=link}