Композиционный генетический код в действии

Пикотехнологические модели четвертичной структуры гистонов.
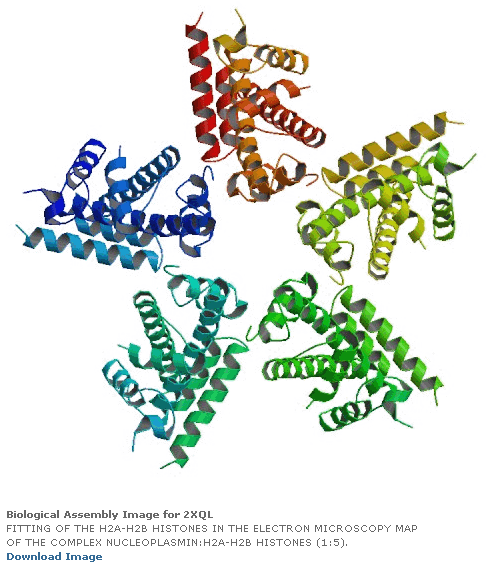
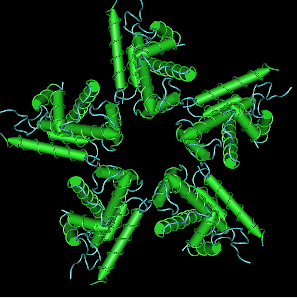


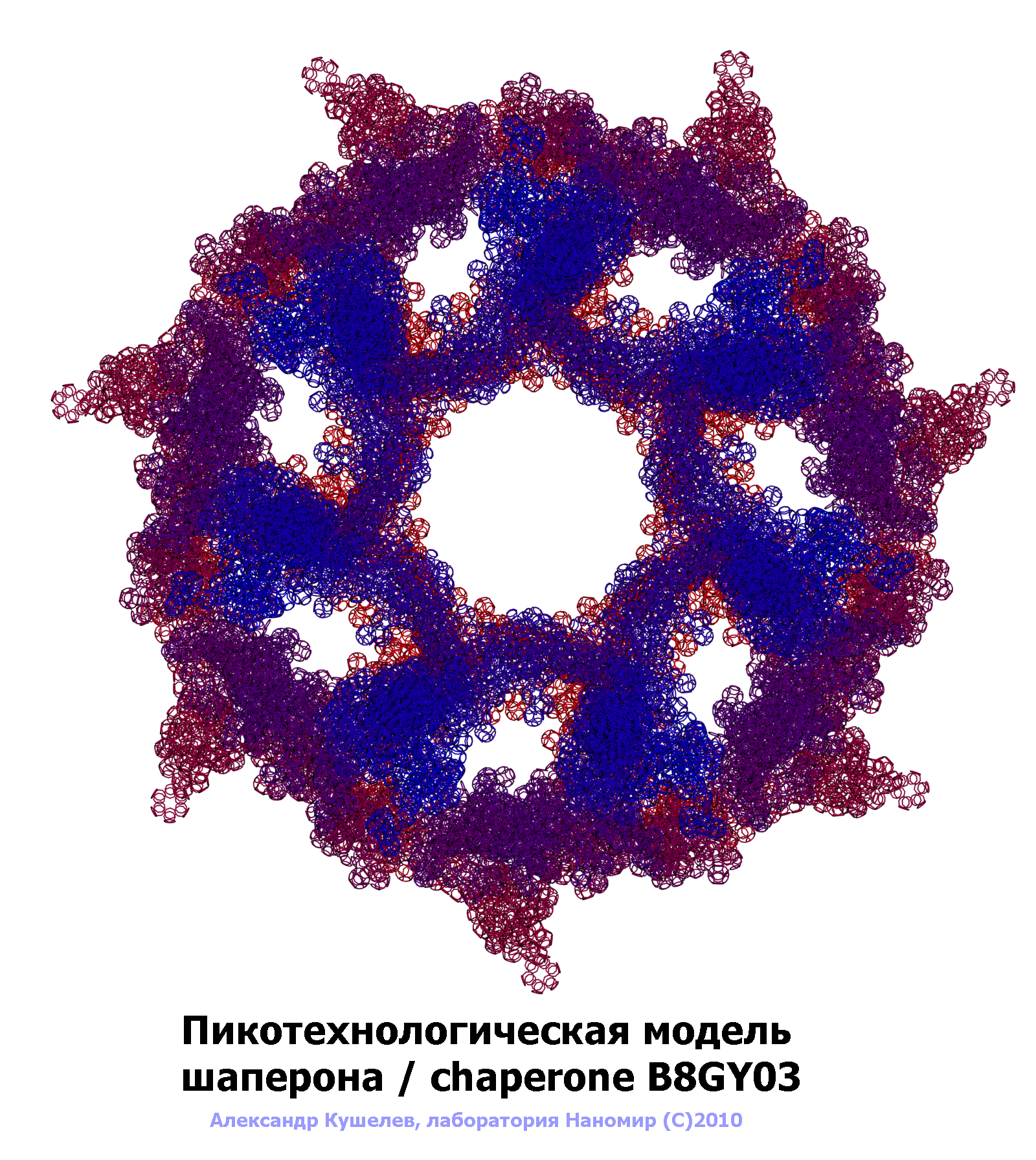
Семиричная симметрия шаперонов.





https://www.nulled.cc/threads/239379/
Александр Кушелев: Ко мне сегодня зашел инвестор, дал 1000 долларов и сказал: Если через неделю сможешь из 1000 долларов сделать 10 000, то я тебе дам миллион. А если не сможешь, то больше не дам ничего. Я его спрашиваю, а если 1000 долларов пропадёт, возвращать надо? Он сказал, что не надо. Я так думаю, что 10 000 долларов из 1000 за неделю сделать нереально, поэтому лучше организовать лотерею. Я отдам эти 1000 долларов случайному человеку сделавшему репост этого сообщения. И не потому что мне не нужны деньги. Просто 1000 долларов для меня погоды не сделает, а ваши репосты дадут мне ... нет, не 10 000 долларов за неделю. Это нереально. Зато позже ваши репосты дадут мне доход от 10 000 долл. в день. Я не боюсь рассказать, как это получится. Дело в том, что это может получиться только у меня. В 1992-ом году я сделал научное открытие, открыл композиционный генетический код. Он отличается от обычного тем, что кодирует не только аминокислоты, но и углы поворота между ними. Казалось бы мелочь, но это позволяет по коду мгновенно определить структуру белка. А пока на определение одной структуры белка уходит до 3 месяцев работы лаборатории рентгеноструктурного анализа. Причём точность новой технологии в ~1000 раз выше. Рентгеном можно определять структуру только кристаллов, а ~97% белков не кристаллизуются. Т.е. моё открытие в 30 раз превосходит рентген! Все лаборатории планеты за один день могут определить 20-30 структур. Моё научное открытие позволяет мне одному определять до 100 структур в день. Это значит, что я могу ежедневно выполнять заказы на миллион евро. Тысячи репостов моего сообщения помогут найти менеджеров, которые будут находить заказы на структуры белка. Мне не жаль отдать им 60% с каждого заказа, например, 6000 евро из 10 000. На таких условиях будут работать тысячи менеджеров, т.к. обычно менеджерам выплачивают несколько процентов от стоимости крупных заказов. Мне выгодно отдать менеджерам больше половины, т.к. я могу делать до 100 заказов в день, поэтому всё-равно заработаю больше. Менеджеров-то много, а я один Осталось придумать правила лотереи, чтобы всё было законно. Я не хочу иметь неприятности. Кроме того, я не люблю обычные лотереи, в которых один выигрывает, а остальные огорчаются. Ведь в этом случае получается, что лотерея даёт радость одному и горе тысячам. И я придумал, как это исправить. Лотерея будет считаться завершенной, если я получу первый коммерческий заказ на структуру белка. Кому логично отдать 1000 долларов? Самому достойному участнику лотереи. Как его определить? Это уже решать вам. Свои 4000 долларов за первую структуру белка я отдам ещё 4 участникам лотереи. Как их определить? Очень просто. В порядке очереди. Кто первый сделал репост, тот первым и получит 1000 долл. Вместе с ним получат ещё 3 участника согласно очереди. Со последующих заказов структур белка я отдаю на выигрыши половину своей доли, т.е. 6000 - менеджеру, 2000 - мне и 2000 - участникам лотереи. С выполнением каждого заказа свои выигрышы будут получать двое из участников в порядке очереди. Если всего участников будет 1000, то после выполнения 500-го заказа каждый из 1000 участников получит по 1000 долл. Как Вам такая лотерея? Верить на слово мне не обязательно. Можно оформить юридически. А чтобы упростить работу менеджеров я готов первые 100 заказов сделать за свой счет. Для меня это - один день работы
Clyde Bardett: I have got the same !!!!!!!!!!!!!!!!!!!!!!
Александр Кушелев: Autotranslation: To me today went investor, gave $ 1000 and said: If in a week can from $ 1000 to do 10 000, then I'll give you a million. And if you can't, then don't give anything. I asked him, and if $ 1000 to fail, the return must be? He said he did not have. I think that 10 000 dollars from 1000 per week impossible to do, so it's better to organize a lottery. I will give these a $ 1,000 casual man made репост of this message. And it's not because I don't need money. Just $ 1000 for me, the weather does not do, and your репосты will give me ... no, not 10 000 USD for the week. It is unreal. But later your репосты will give me an income of $ 10,000 a day. I'm not afraid to tell the story of how this will work out. The fact that it can happen only to me. In 1992 I made a scientific discovery, opened composite genetic code. It differs from a standard that encodes not only amino acids, but and angles between them. It would seem a trifle, but it allows the code instantly determine the structure of the protein. Meanwhile, the definition of one of protein structure takes up to 3 months the work of the laboratory of x-ray analysis. The accuracy of new technologies in about 1000 times higher. X-ray you can define the structure of only the crystals, and AC 97% of proteins do not crystallize. I.e. my discovery in 30 times greater than the x-ray! All laboratories of the planet in one day can determine 20-30 structures. My scientific discovery allows me to one define up to 100 structures in the day. This means, that I may daily perform orders for one million euros. Thousands of репостов of my posts will help you to find managers who will find orders for protein structure. I'm not sorry to give them 60% with each order, for example, 6,000 euros from 10 000. In such conditions they will work thousands of managers, because it usually managers pay a few percent of the value of major orders. Is it profitable for me to give managers more than half, because I can make up to 100 orders a day, so all the same to earn more. Managers much, and I'm one Remains think of the rules of the lottery, so that everything was legal. I don't want to have troubles. In addition, I do not like conventional lottery, in which one wins, and the rest of the people will be annoyed. In fact, in this case it turns out that the lottery gives joy to one and mount the thousands. And I figured out how to fix it. The draw will be considered completed, if I get the first commercial order for the structure of protein. To whom it is logical to give $ 1000? Most worthy participant of the lottery. How to define it? It is already up to you. Its $ 4000 for the first structure of protein I will give another 4 participants of the lottery. How to define them? Very simply. In turn. Who first made репост, the first and will receive $ 1,000. Together with him will receive 3 more of the participant according to the queue. With subsequent orders structures of protein I give the winnings half of its shares, i.e. 6000 - Manager of the 2 - me and 2 - participants of the lottery. With the execution of each order your prize will receive two of the participants in the line. If all of the participants will be 1000, after the 500-th order each of the 1000 participants will receive $ 1,000. As You are such a lottery? Believe the word I don't have to. Can be issued legally. And in order to simplify the work of the managers I am ready to the first 100 orders to do at his own expense. For me it's one day of work.
2012.04.07 13:44:30
Пикотехнология - новый подход в моделировании пространственной структуры белка. Кушелев А.Ю. Соколик В.В.
Текстовый вариант статьи:
ПИКОТЕХНОЛОГИЯ – НОВЫЙ ПОДХОД В МОДЕЛИРОВАНИИ
ПРОСТРАНСТВЕННОЙ СТРУКТУРЫ БЕЛКА
Кушелев А.Ю., Соколик В.В.
Научно-исследовательская лаборатория "Наномир", Дмитров
ГУ «Институт неврологии, психиатрии и наркологии АМНУ», Харьков
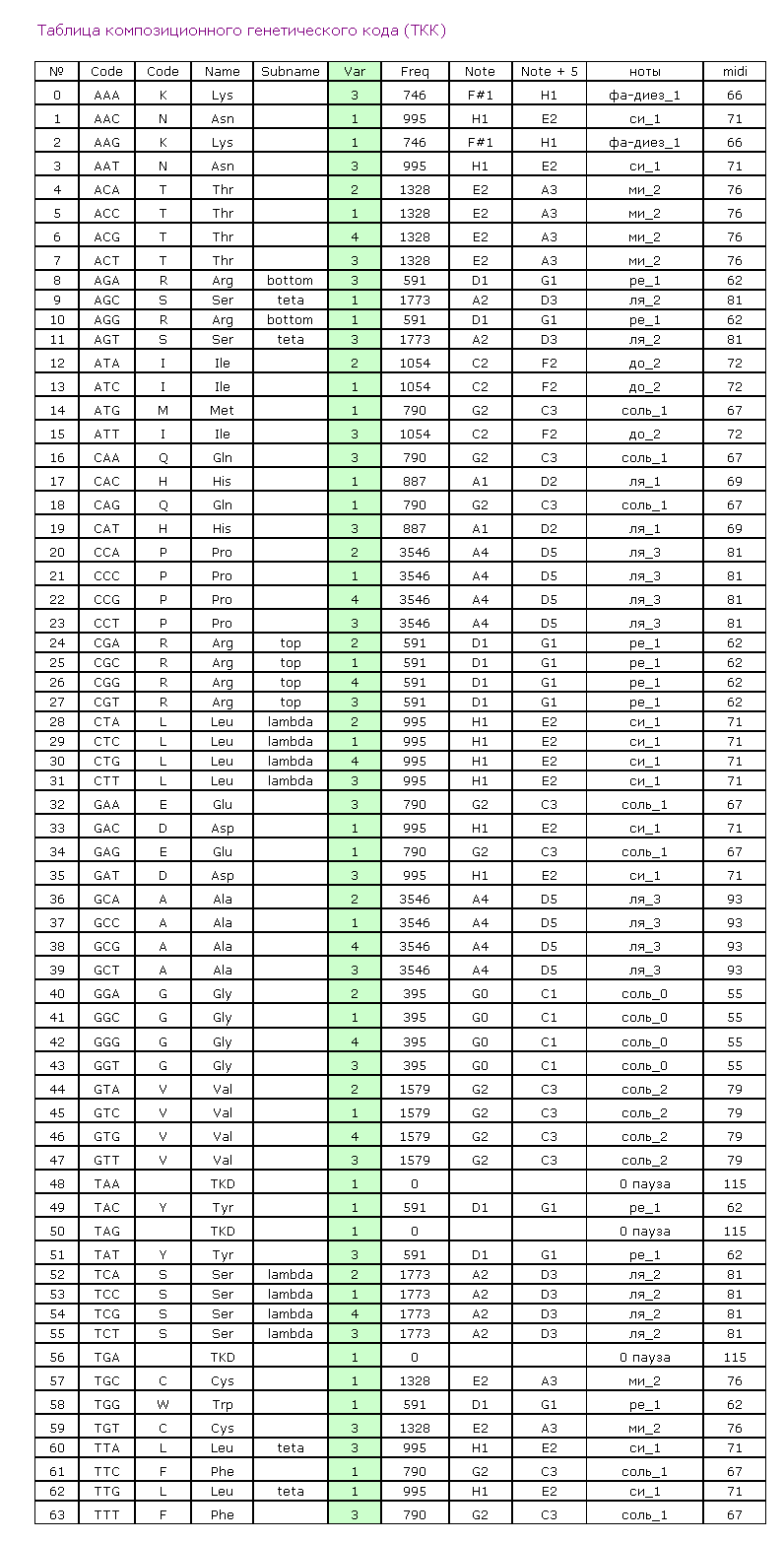
В настоящее время наибольшей популярностью пользуются научные исследования с приставкой «нано»: речь идёт об объектах, размер которых лежит в диапазоне 1-100 нанометров [3, с. 28; 16, c. 6]. Однако при моделировании атомов в составе биомолекул возникла необходимость оперировать конфигурацией электронов, формирующих внешний валентный уровень, который и определяет объем, занимаемый тем или иным атомом в пространстве, а это уже размерность пикометров (10-12 м). Пикотехнология, методический подход для моделирования пространственной структуры белка, опирается на следующие представления о микромире:
· кольцегранная структура электронных орбиталей (электронов) в атоме, которая детерминирует в первом приближении форму атома в виде усечённого октаэдра (рис. 1), грани которого – электроны валентного энергетического уровня [17, c. 49; 4, с. 46; 1, с. 73];
· геометрический алгоритм объединения атомов в молекулах определяет не только межатомные расстояния (длины связей), но прежде всего углы поворота по этим связям, кратные 120о в трёхмерном пространстве [2, c. 237; 5, с. 137];
· ротамерный вариант пептидной связи, которая объединяет аминокислотные остатки в полипептид, детерминирован третьим нуклеотидом кодона [2, c. 237; 18, c. 347] и реализуется в ходе матричного синтеза структурного шаблона белка на рибосоме пулом изоакцепторных тРНК [6, c. 13].

А Б
Рис. 1. Кольцегранная модель валентной электронной оболочки атома из 8 электронов (А) и её аппроксимация усеченным октаэдром (Б) в моделях белков.
Десять лет назад А.Ю. Кушелевым была сформулирована идея композиционного генетического кода [2, c. 237], которая была расширена представлениями о кодировании ротамерии пептидной связи и структурного шаблона белка [18, c. 348; 19, с. 275; 20, c. 118]. В таблице композиционного генетического кода каждому варианту композиционного кода, в зависимости от кодона, соответствовало определённое значение композиционного угла, под которым в ходе матричного синтеза происходит присоединение очередного аминокислотного остатка к растущей полипептидной цепи. Данная таблица легла в основу алгоритма первого варианта компьютерной программы «Пикотехнология» для моделирования пространственной структуры белка по детерминирующей его нуклеотидной последовательности. Дальнейшая разработка данной проблемы привела к уточнению не только самой таблицы, но и к введению таких понятий, как ротамерия пептидной связи и структурный шаблон белка. Было установлено, что в геноме третьим нуклеотидом кодона детерминирован один из трёх ротамерных вариантов пептидной связи, которым аминокислотный остаток (закодированный дуплетом первых двух нуклеотидом кодона) присоединяется к растущей полипептидной цепи (табл. 1).
Таблица 1. Генетический код структурного шаблона белка

XYZ – первый, второй и третий нуклеотиды в кодоне; R, 0, L – ротамерные варианты пептидной связи (РВПС); uaa, uag, uga – Stop-кодоны.
Ротамерные варианты пептидной связи различаются между собой углом поворота по оси пептидной связи ?, кратным 120о. Крайне важно понимать, что ротамерный вариант пептидной связи реализуется в процессе синтеза белка в рибосоме и дальнейшее вращение по уже образовавшейся полуторной пептидной связи становиться невозможным. Поэтому с рибосомы сходит совершенно индивидуальный структурный шаблон белка из последовательности ротамерных вариантов пептидной связи, в соответствие с информацией, содержащейся в его гене. Именно этим обстоятельством мы объясняем невозможность синтезировать нематричным способом функционально активные большие молекулы белковых ферментов или рецепторов. Твердофазный синтез реализован только для небольших неструктурированных пептидов, которые характеризуются избыточной конформационной подвижностью, в силу чего их функциональные конформации определяются взаимодействием с белками-партнёрами в составе гетерокомплексов, а не индивидуальным структурным шаблоном [13, с. 38].
Для конформеров вторичной структуры белка характерна периодичность, поэтому, правая спираль в структурном шаблоне белка кодируется последовательностью кодонов с С/G в третьей позиции, ?-тяж – повторением кодонов с А, а левая спираль – последовательностью кодонов с Т в третьем положении (табл. 2). Неструктурированные фрагменты кодируются чередованием кодонов с С/G, А и Т в третьей позиции.
Таблица 2. Кодирование конформеров вторичной структуры белка

У R, 0 и L-ротамеров все атомы пептидной группы (C? (i), C (i), O (i), N (i+1) H (i+1)) компланарны, кроме C? (i+1). C? (i+1) атом каждого аминокислотного остатка не принадлежит плоскости пептидной группы, благодаря чему происходит сворачивание полипептидной цепи в конформеры вторичной структуры белка (правая ?-спираль, ?-тяж, левая 3/10-спираль) ещё в рибосоме (рис. 2), а не после синтеза полипептидной цепи из практически единственного транс-изомера (цис-изомер только у пролина) в виде плоской ленты, сворачивание которой достигается поворотами на углы ? и ? по связям СО—C? и NH—C?, как предполагали ранее [9, с. 134]. Пластичность структурного шаблона в ходе посттрансляционного фолдинга и конформационная подвижность белка при взаимодействии с лигандами достигаются единственно возможным поворотом по оси связи NH—C? на приращение угла ?, кратное 120о [5, с. 139].

Рис. 2. Схема образования правого (R), нулевого (0) и левого (L) ротамерных вариантов пептидной связи.
Данный механизм трансляции генетической информации является эволюционно новым. Его формирование у эукариот было обусловлено необходимостью синтеза больших и сложных белков в виде структурного шаблона, максимально приближенного к функциональной конформации этих белков, чтобы их фолдинг имел наибольшие скорость и КПД. У прокариот и органелл эукариот (митохондрии, хлоропласты) третий нуклеотид кодонов в генах небольших полипептидов ещё не является информационным, поэтому на нём и наблюдается воблирование по описанному Ф. Криком механизму [12, с. 368]. Это обусловлено отсутствием пула изоакцепторных тРНК с модифицированными нуклеотидами в первом положении антикодона [6, с. 11] и нередко отсутствием филогенетически более молодых областей в структуре тРНК [15, с. 6730], т.е. недоразвитием звена, реализующего информацию третьего нуклеотида.
Выше изложенные положения легли в основу алгоритма компьютерных программ Secondary Structure Protein (SSP) и Three-dimension Structure Protein (TSP), которые по нуклеотидной последовательности мРНК позволяют смоделировать схему вторичной структуры и визуализировать индивидуальный структурный шаблон любого белка. Эту первичную информацию о белке можно использовать в дальнейшем моделировании фолдинга функциональной конформации белка с учетом физ-химии его микроокружения, посттрансляционных модификаций, взаимодействия с лигандами методами молекулярной динамики наравне с информацией о наиболее стабильном конформере, которую извлекают из рентгенограмм кристаллов белков. Преимущество данного подхода состоит в возможности быстрого моделирования индивидуальной пространственной структуры отдельной молекулы любого белка (даже если он не кристаллизуется, и никогда не сворачивается, как, например, регулятор клеточного деления Sic1 [11, с. 152]) с точностью до электрона (пикотехнология), опираясь лишь на информацию о нём в геноме. То есть, мы in silico воспроизводим трансляцию генетической информации в индивидуальный структурный шаблон белка, а не занимаемся поиском самой стабильной или «быстро достигаемой» устойчивой его конформации из 10100 возможных [14, с. 44], как это происходит при конформационном анализе поверхности потенциальной энергии молекулы белка громоздкими методами систематического поиска, Монте-Карло или молекулярной динамики с целым рядом ограничений и приближений [10, с. 41]. Не исключено, что большинство белков именно из конформации своего структурного шаблона максимально быстро, а главное однозначно, фолдируют в нативную конформацию с минимумом свободной энергии, формируя, таким образом, «устойчивое большинство» конформационно лабильного белкового пула.

Рис. 3. Пикотехнологические модели структурных шаблонов некоторых белков и их комплексов.
В последнее время А.С. Спирин, отклоняясь от постулата о матричном синтезе белка в виде развёрнутой полипептидной цепочки [7, с. 5], предположил, что в самой рибосоме полипептид синтезируется сразу в виде ?-спирали и по желобу выталкивается наружу по мере трансляции мРНК [8, с. 437]. Этой прогрессивной гипотезе, которая основывается на подавляющем (74%) большинстве «спиральных» кодонов в генах эукариот, остался один шаг до представления о трансляции структурного шаблона белка не только в виде ?-спирали.
С помощью пикотехнологии были декодированы и смоделированы структурные шаблоны более 100 белков (рис. 3), сопоставительный анализ которых с экспериментальными данными Protein Data Bank (PDB) позволил подтвердить предположение о генетически закодированных размерах и местоположении конформеров вторичной структуры в нативной конформации этих белков [18, c. 348].
Итак, пикотехнология – это современный, точный и удобный методологический подход в арсенале молекулярной биологии для моделирования пространственной структуры белков, исходя из той информации генома о них, которой располагает сама клетка.
Список литературы
1. Кушелев А., Полищук С., Писаржевский С. Формы, механизмы, энергия наномира: Доступна ли энергия эфира для космических полётов? // Электроника: Наука, Технология, Бизнес. – 2002. – № 6. – С.72–76.
2. Кушелев А.Ю., Полищук С.Е., Неделько Е.В. и др. Построение масштабной модели структуры белка // Актуальные проблемы современной науки. – 2002. – № 2. – С. 236–243.
3. Нанонаука и нанотехнологии: энциклопедия систем жизнеобеспечения / Моск. гос. техн. ун-т им. Н. Э. Баумана; ред. О. О. Аваделькарим; гл. ред.: Чуньли Бай, С. П. Капица. — М.: Магистр-Пресс : Изд-во ЮНЕСКО : EOLSS, 2009. — 991 с.
4. Огжевальский З.И. 1972. Пространственные модели атомов, молекул и кристаллов. Москва, 1972. – 118 c.
5. Соколик В.В. Карта Рамачандрана: ротамерия пептидной связи и фолдинг белка // VII Международная научно-техническая конференция «Актуальный вопросы биологической физики и химии». БФФХ-2011, Севастополь. – 2011. – С.137–139.
6. Соколик В.В. Загадка изоакцепторных тРНК // II Всероссийская Интернет-Конференция «Актуальные проблемы биохимии и бионанотехнологии», Казань. – 2011. – С. 11-15.
7. Спирин А.С. Биосинтез белка: элонгация полипептида и терминация трансляции // Соросовский образовательный журнал. – 1999. – № 6. – С. 2–7.
8. Спирин А.С. Молекулярная биология: рибосомы и синтез белка. – М: «Академия», 2011. – 496 с.
9. Финкельштейн А.В., Птицын О.Б. Физика белка. - М.: Книжный дом «Университет», 2002. – 298 с.
10. Хёльтье Х.-Д., Зиппль В., Роньян Д., Фолькерс Г. Молекулярное моделирование. М.: БИНОМ. Лаборатория знаний, 2010. – 320 с.
11. Chouard T. Structural biology: Breaking the protein rules // Nature. – 2011. – V. 471, № 7337. – P. 151–153.
12. Crick F.H.C. The Origin of the Genetic Code // J. Mol. Biol. – 1968. – V. 38. – P. 367–379.
13. Fink A.L. Natively unfolded proteins // Curr. Opin. Struct. Biol. – 2005. – V.14, № 1. – P. 35-41.
14. Levinthal C. Are there pathways for protein folding // J. Chim. Phys. – 1968. – V. 65. – P. 44–45.
15. Maizels N., Weiner A.M. Phylogeny from function: Evidence from the molecular fossil record that tRNA originated in replication, not translation. // Proc.Nat.Acad.Sci.USA. – 1994. – V. 91, № 15. – P. 6729–6734.
16. Ratner M., Ratner D. Nanotechnology: a gentle introduction to the next big idea, 2003. – 195 p.
17. Snelson K. A design for the atom // Industrial design. – 1963. – № 1. – P. 48–57.
18. Sokolik V.V. Protein is coded in genome and synthesized in ribosomes as a structural template of a rotameric version sequence of peptide bound configuration // The International Moscow Conference on Computational Molecular Biology, МССМВ-11, Moscow. – 2011. – P. 347–348.
19. Sokolik V.V. Modeling of the individual structural template of protein on determining it nucleotide sequences // VII Международная конференция по биоинформатике, регуляции и структуры геномов и системной биологии. BGRS\SB-2010, Новосибирск. – 2010. – С. 275.
20. Sokolik V.V. Algorithm of protein structural template decoding according to its determined nucleotide sequence // Fist International Conference “Fundamental medicine: From scalpel toward Genome, Proteome and Lipidome”, Pax Grid Virtual Conferences, Kazan. – 2011. – P. 117–119.
Подписи под иллюстрациями.
Рис. 3: Гистоновый комплекс 5 H2A + Н2В_DROME (P84051, P02283)
Шаперон A0KFQ4_AERHH (A0KFQ4)
Суперспираль коллагена CO2A1_HUMAN (Р02458)
Белковый комплекс ?-протеина с ?-тубулином TAU_HUMAN + TBA1A_HUMAN
Инсулин INS_HUMAN (Р01308)
Лизоцим LYSC_CHICK (P00698)
Рибонуклеаза Н1 RNH_ECOLI (P0A7Y4)
Аполипопротеин Е APOE_HUMAN (P02649)
Гемоглобин (А-цепь) HBA_HUMAN (P69905)